Ювенильді-гормондық эстераза - Juvenile-hormone esterase
| ювенильді гормонның эстеразасы | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 | |||||||||
| Идентификаторлар | |||||||||
| EC нөмірі | 3.1.1.59 | ||||||||
| CAS нөмірі | 50812-15-2 | ||||||||
| Мәліметтер базасы | |||||||||
| IntEnz | IntEnz көрінісі | ||||||||
| БРЕНДА | BRENDA жазбасы | ||||||||
| ExPASy | NiceZyme көрінісі | ||||||||
| KEGG | KEGG кірісі | ||||||||
| MetaCyc | метаболизм жолы | ||||||||
| PRIAM | профиль | ||||||||
| PDB құрылымдар | RCSB PDB PDBe PDBsum | ||||||||
| Ген онтологиясы | AmiGO / QuickGO | ||||||||
| |||||||||
Жылы энзимология, ювенильді гормон эстеразасы (JH этеразасы) болып табылады фермент бұл катализдейді The гидролиз туралы кәмелетке толмаған гормон. Мысалы, кәмелетке толмаған гормон II (табылған Лепидоптера ):
- метил (2E, 6E) - (10R, 11S) -10,11-эпоксид-3,7,11-триметилтридека-2,6-диеноат + H2O (2E, 6E) - (10R, 11S) -10,11-эпоксид-3,7,11-триметилтридека-2,6-диеной қышқылы + метанол.

Номенклатурасы және қызметі
Бұл фермент тұқымдасына жатады гидролазалар, атап айтқанда, карбоксилмен жұмыс жасайтындар күрделі эфир облигациялар The жүйелік атауы осы фермент класының метил- (2E,6E)-(10R,11S) -10,11-эпоксид-3,7,11-триметилтридека-2,6-диеноат ацилгидролаза. Жалпы қолданыстағы басқа атауларға JH эстераза, ювенильді гормон эстеразы және юбильді гормон карбоксиэстераза жатады.
Ювеналды гормон (JH) жәндіктерді басқарады метаморфоз.[1] JH титрінің төмендеуі қуыршақ кезегін бастаса, жоғары JH титрлары личинка күйін сақтайды [2] қуыршақ сатысында қуыршақ тіндеріне личинка тіндерінің синтезінен алыс тіндердің міндеттемелерінің өзгеруі.[3] Lepidoptera-дағы соңғы личинка жасушасының басында JH титрінің төмендеуі метаболизмнің жоғарылауымен байланысты [4] және синтездің төмендеуі.[5] Ішінде Лепидоптера, JH бастапқыда эфир гидролизімен метаболизденеді;[6] гидролиздеуге қабілетті эфираздарды анықтайды гемолимф кейде JH титрінің төмендеуімен сәйкес келетін соңғы дернәсілдер кезінде.[7] JHE-лері табиғи түрде пайда болған JH металдарының 2E метил эфирі үшін де селективті болып табылады.[8] Бұл зерттеулер JHE-дің JH титрін реттеуде маңызды болуы мүмкін, сондықтан қуыршақ сатысының басталуы мен міндеттемесіне қатысуы мүмкін. JHE-ді өндіретін көрінеді семіз дене [9] және бұл өндірісті экзогендік JH арқылы ынталандыруға болады Гиалофора қуыршақ, JHE белсенділігінен айрылған кезең.[10] Жақында JH-дің JHE белсенділігін ынталандыру ересектерде де байқалды Leptinotarsa decemlineata [11] және қуыршақтары Galleria mellonella.[12] Алайда, осы уақытқа дейін, осы ферменттер бірінші кезектегі маңызы бар деп саналатын соңғы личинка кезінде осы құбылысты бірде-бір зерттеулер зерттеген жоқ. Осылайша, зертхана зертханалық тексерісті өз мойнына алды гемолимф Қырыққабат луперінің соңғы дернәсілдік кезіндегі JHE реттелуі, Trichoplusia ni.
JH эстераза индукциясы
Ювеналды гормон эстераза жәндіктердің басында кездесетін факторлар әсерінен туындайды.[13] Сонымен қатар, ол жәндіктерді табиғи жолмен өңдеу арқылы шақырылады кәмелетке толмаған гормон, JH I ең күшті индуктор.[14] JH синтетикалық агонистері дәл осындай белсенділікті JH I-ге қарағанда төмен потенциалда болса да дәлелдеді.[15] Басқа зерттеуде жәндіктердің басында болатын факторлар JH белсенділігінің индукторлары екендігі дәлелденді.[16] Лепидоптеран личинкаларының ашығуы JH эстеразасының пайда болуын да тудырады.[17]
JH этераза ингибиторлары
JH эстеразасының ингибиторлары болып табылатын бірқатар қосылыстар табылды. Олардың көпшілігі инсектицидтер болып табылады, олар екі негізгі құрылымдық топқа, яғни фосфоамидотиолаттар мен S-фенилфосфаттарға; карбамат инсектицидтер де сыналды.[18] Әдетте ең күшті ингибиторы IC бар этоксиотиофенилфосфамидотиолат болды.50 <1 нМ. Бұл зерттеуге ерекше қызығушылық бар этил және изопропил табиғи JH аналогтары этеразамен бөлінбеді, бұл оның бар екендігін көрсетеді метил арнайы эфирге. JH I және JH III 5 мкМ номиналды концентрациясында сыналды. Кейінірек трифторометил кетон (3-октилтио-1,1,1-трифлуоро-2-пропанон) Trichoplusia ni-нің JH эстеразасын баяу, тығыз байланыстыратын ингибиторы, дәл сол лепидоптеран болып шықты. осы бөлімдегі басқа зерттеуде.[19] Бұл зерттеуде бұл қосылыстың тежелуіне өте күрделі кинетикалық анализдер берілген (OTFP аббревиатурасы), JH I ферменттің әсерінен JH III-ге қарағанда тез ыдырайды, ал Km мәні JH III-тен екі есе артық.
JH этеразасының уақытқа байланысты ауытқуы және жәндіктердің дамуына байланысты
JH эстеразасы және JH эпоксид гидролазы JH әсерін тоқтатуда өте маңызды. Жасөспірімдер гормонын байланыстыратын ақуыздардың рөлі де маңызды, өйткені олар мүмкіндік береді кәмелетке толмаған гормон гидролитикалық ферменттерден қорғау.[20] Бұл өте күрделі сценарийді құрайды, оны зерттеу қиын, сонымен қатар әртүрлі түрлерді ажырату қиын. JH, JH қышқылы, экдизон және JH титрлары бесінші дернәсілдік стадион кезінде даму функциясы ретінде Manduca sexta-ның дәл уақытылы дернәсілдерінде жасалды. Бұл дернәсілдерде JH принципі JH I және JH II, төмен деңгейлері JH 0 және JH III. Төртінші стадионның соңында JH I және II үлкен шыңы бар, олардың қышқыл метаболиттерінің төменгі деңгейлерімен бірге жүреді. Содан кейін JH эстеразасының кең шыңы 1,5-тен 4-ші күнге дейін басталады. Кейіннен экдистероидтық титрлер 3,5-ші күні аздап көтеріледі, содан кейін 5-ші күннен басталып, 5-ші күннен төмен деңгейлерде жалғасатын эксидероидтық титрлер бұл күрт жүреді. JH I және JH II шыңы, 4-ші күннен басталып, 6-шы күні аяқталады. JH I қышқыл титрлері JH I титрімен бірдей, тек 7-ші күнді қоспағанда, тек JH I қышқылының шыңы. Бұл экдистероидтық титрлардың төмендеуі сияқты.[21] JH этераза шыңдарының уақыты өте ұқсас және экдизон байқалды Galleria mellonella.[22] Бұл мәліметтер лепидоптерандар үшін классикалық модельге сәйкес келеді, онда әр личинка балқымасында JH жоғары, бірақ бірге көтерілуі керек экдизон қуыршақтан бұрын қуыршақ бальтасын бастайды. Олар корпорация allata экстракорпоралды 0-ші күнде жүргізетін басқалар ұсынған модельге сәйкес келеді M. sexta дернәсілдер жоғары деңгейдегі JH бөледі, бірақ 4-ші тәулікте тек JH қышқылын өндіруге ауысады, содан кейін JH шыңын жасау үшін қиял дискілері арқылы метилденеді.[23] Алайда, CA шығарған JH салыстырмалы мөлшерінің секрециясы Manduca sexta in vivo титрлерінен айтарлықтай ерекшеленетіні анықталды.[24] JH титрлерін зерттеу Trichoplusia ni уақытша және эдизон секрециясына қатысты JH импульсінің уақыты туралы өте ұқсас тұжырымдар жасады. Алайда бұл түрдегі JH принципі JH II болып табылады. Эстераза ингибиторы, EPPAT инъекциясының күшейгені анықталды кәмелетке толмаған гормон титрлер, ал аштық жасөспірімдер гормонының титрін жоғарылататыны анықталды. Сонымен қатар, личинкалардың паразиттелуі Хелонус sp. (Hymenoptera) JH II титрін төмендететіні анықталды, бірақ паразиттен алынған JH III титрінің жоғарылауын тудырды.[25]
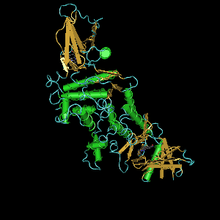
JH эстераза ақуызының құрылымы
Темекі мүйізденетін эстеразаның кристалдық құрылымы Manduca sexta 3-октилтио-1,1,1-трифторопропан-2-бір (OTFP) аналогты ингибиторымен белсенді учаскеге кешенді түрде белсенді учаскемен байланысқан кешенді түрде шешілді. Бұл кристалды құрылымда ұзын, гидрофобты байланыстырушы қалта бар, соңында еріткіш жетпейтін каталитикалық триада бар. Құрылым JHE және оның субстраттары мен ингибиторлары арасындағы байқалған көптеген өзара әрекеттесулерді түсіндіреді, мысалы, метил эфирлерге қарсы этил немесе изопропил күрделі эфирлер және гидрофобты магистральдар.[26] Фермент өте тиімді, мысық / км кем дегенде 3 x 107 M-1 s-1 құрайды. Бастапқы тізбектің ұзындығы 583 амин қышқылынан тұрады, 22 аминқышқыл ақуызынан тұрады. Белсенді формадағы есептелген Mr - 62,1 кДа.
Жазбаша әдебиеттер
Абдель-Аал, Ю.А.И., Ро, Р., Хаммок, Б.Д., 1984. Екі трифлуорометилкетон және О-этил, S-фенилфосфорамидотиоат арқылы тежелетін ювенильді гормон эстеразасының кинетикалық қасиеттері. Зиянкестер. Биохимия. Физиол. 21, 232-241.
Бейкер, Ф.С., Цай, Л.В., Ройтер, КС, Шули, Д.А., 1987. Бесінші стадион Manduka sexta-ны дамыту кезінде inHiv in Vivo-да JH, JH қышқылы және экдистероид титрі және JH эстераза белсенділігі. Жәндіктер биохимиясы. 17, 989-996.
Браун, Р.П., Уайатт, Г.Р., 1995. Ересек шегірткелердегі еркектердің қосымша безінің өсуі: Ювенильді гормонның рөлі, JH эстеразы және JH байланыстыратын ақуыздар. Арх.Жәндіктер биохимиясы.Физиол. 30, 383-400.
De Kort, C.A.D., Granger, N.A., 1996. JH титрлерінің реттелуі: деградациялық ферменттер мен байланысқан ақуыздардың өзектілігі. Арка. Жәндіктер биохимиясы. Физиол. 33, 1-26.
Гилберт, Л.И., Гудман, В., Болленбахер, В.Е., 1977. Жәндіктердегі реттеуші липидтер мен стеролдардың биохимиясы., Гудвин, Т.В. (Ред.), Липидтер биохимиясы II. Биохимияның халықаралық шолуы. University Park Press, Балтимор, I-50 б.
Хаммок, Б., Новок, Дж., Гудман, В., Стамудис, В., Гилберт, Л.И., 1975. Гемолимфа байланыстыратын протеиннің ювенильді-гормондық тұрақтылыққа және таралуына Мандука-Секста май-денесі мен қиял дискілеріндегі әсері Invitro . Молекулалық және жасушалық эндокринология 3, 167-184.
Хаммак, Б.Д., Квистад, Г.Б., 1976. Ювеноидтардың жәндіктермен деградациялық метаболизмі, ішінде: Гилберт, Л.И. (Ред.), Ювеналды гормондар. Пленум Пресс, Нью-Йорк, 374–393 бб.
Хаммок, Б.Д., Спаркс, ТК, Мумби, СМ, 1977. Таракан гемолимфасынан JH эстеразаларының селективті тежелуі. Зиянкестер. Биохимия. Физиол. 7, 517-530.
Хвансу, К., Редди, Г., Кумаран, А.К., Болленбахер, Ве, Гилберт, Л.И., 1979. Галлериа-Меллонелладағы ювенильді гормондардың эстераза-белсенділігі, экдизон титрі және жасушалық қайта бағдарламалау арасындағы өзара байланыс. J. жәндіктер физиолы. 25, 105-111.
Джонс, Г., Ханзлик, Т., Хаммок, Б.Д., Шули, Д.А., Миллер, Ч.А., Цай, Л.В., Бейкер, Ф.К., 1990. Трихоплусия нидің соңғы және соңғы литальды стадионындағы жасөспірім гормоны титрі. J. жәндіктер физиолы. 36, 77-83.
Джонс, Г., Уинг, К.Д., Джонс, Д., Хаммак, Б.Д., 1980. Қырыққабат луперінің құртындағы ювенильді гормон эстеразасын реттейтін бас факторлардың көзі және әрекеті, Trichoplusia ni. J. жәндіктер физиолы. 27, 85-91.
Крамер, С.Ж., 1978. Колорадо қоңызы, Leptinotarsa decemlineata-да JH-ге тән эстеразалардың белсенділігін реттеу. J. жәндіктер физиолы. 24, 743-747.
Ниджут, Х., Уильямс, С., 1974. Темекі мүйізіндегі мюльтингті және метаморфозды бақылау, Мандука Секста (Л.): жасөспірім гормонының секрециясын қуыршақ үшін қоздырғыш ретінде тоқтату J. Exp. Биол. 61, 493-450.
Nijhout, H.F., 1975. Темекі мүйізі құртының личинкаларындағы жасөспірімдер гормонының әсер ету динамикасы. Теңіз биологиялық зертханасының биологиялық бюллетені, Woods Hole 149, 568-579.
Nowock, J., GILBERT, L., 1976a. Жәндіктердің жасөспірім гормонының титрін реттейтін факторларды in vitro талдау, Курстак, Э., Мараморош, К. (Ред.), Омыртқасыздар тіндерінің мәдениеті. Academic Press, Нью-Йорк, 203–212 бет.
Новок, Дж., Гилберт, Л.И., 1976б. Жәндіктердің ювенильді гормонының титрін реттейтін факторларды in vitro талдау, мыналарда: K., K.E.a.M. (Ред.), Омыртқасыздардың тіндік мәдениеті. Academic Press, Нью-Йорк, 203–212 бет.
Плапп, Ф.В., кіші, Кариньо, Ф.А., Вей, В.К., 1998. Үйден ұшатын ақуызды байланыстыратын ювенильді гормон және оның инсектицидтерге төзімділігімен байланысы. Арка. Жәндіктер биохимиясы. Физиол. 37, 64-72.
Прествич, Г.Д., Войтасек, Х., Ленц, А.Ж., Рабинович, Дж.М., 1996. Жасөспірім гормондарын байланыстыратын және метаболиздейтін ақуыздардың биохимиясы. Арка. Жәндіктер биохимиясы. Физиол. 32, 407-419.
Редди, Г., Хвангсу, К., Кумаран, А.К., 1979. Балауыз көбелегіндегі жасөспірімдер гормонының эстераза-белсенділігіне әсер ететін факторлар, Галлерия-Меллонелла. J. жәндіктер физиолы. 25, 65-71.
Риддифорд, Л.М., 1976. in vitro жағдайында жәндіктердің эпидермиялық жасушаларының міндеттемелерін гормоналды бақылау. Табиғат 259, 115-117.
Санбург, Л.Л., Крамер, К.Ж., Кезди, Ф.Ж., Заң, Дж.Х., 1975а. Мандука секста темекі мүйізді гемолимфасындағы жасөспірім гормонына тән эстеразалар. J. жәндіктер физиолы. 21, 873-887.
Санбург, Л.Л., Крамер, К.Ж., Кезди, Ф.Ж., Заң, Дж.Х., Оберландер, Х., 1975б. Ювенильді гормон эстеразаларының және тасымалдаушы ақуыздардың жәндіктердің дамуындағы рөлі. Табиғат 253, 266-267.
Слэйд, М., Зибитт, С.Х., 1972. Жәндіктердегі және сүтқоректілердегі цекропияның ювенильді гормонының метаболизмі, Менн, Дж. Дж., Бероза, М. (Eds.), Жәндіктердің кәмелетке толмаған гормондары: химия және әрекет. Academic Press, Нью-Йорк, 155–176 бб.
Sparagana, SP, Bhaskaran, G., Barrera, P., 1985. Manduca sexta prepupae-дің жасанды гормонының метилтрансфераза белсенділігі. Арка. Жәндіктер биохимиясы. Физиол. 2, 191-202.
Спарагана, С.П., Бхаскаран, Г., Дахм, К.Х., Реддл, В., 1984. Кәмелетке толмаған гормондардың өндірісі, кәмелетке толмаған гормондардың эстеразы және ювенильді гормон қышқылы метилтрансфераза Manduca sexta. J. Exp. Zool. 230, 309-313.
Sparks, T.C., Hammock, BD, 1979. Қырыққабат луперінің соңғы дернәсілдік кезіндегі жасөспірім гормондарының эстеразаларын индукциялау және реттеу, Trichoplusia ni. J. жәндіктер физиолы. 25, 551-560.
Sparks, TC, Hammock, BD, Riddiford, LM, 1983. Manduca sexta (L.) Гемолифтік ювенильді гормон эстеразасы - ингибирлеу және реттеу. Жәндіктер биохимиясы. 13, 529-541.
Sparks, T.C., Wing, KD, Hammock, BD, 1979. Трихоплусия нидегі жәндіктер жасөспірімдер гормонының эстеразасының индукциясына анти-гормон-гормон гормонының ЕТБ имитациясы. Life Sci. 25, 445-450.
Винс, Р.К., Гилберт, Л.И., 1977. Жасөспірім гормонының эстераза белсенділігі дәл уақытында дерлік дерлік дернәсілдер мен Manduca sexta фарат қуыршақтарында. Жәндіктер биохимиясы. 7, 115-120.
Вейрич, Г., Рен, Дж., 1973. Мандука секста гемолимфасынан алынған ювенильді гормон эстеразасының субстрат ерекшелігі. Life Sci. 13, 213-226.
Вейрих, Г.Ф., Рен, Дж., 1976. Жәндіктердің дамуындағы ювенильді-гормондық этераза: салыстырмалы зерттеу. Физиологиялық зоология 49, 341-350.
Уитмор, Д., Гилберт, Л.И., Итчер.Пи, 1974. Гемолимф карбоксилестеразаларының шығу тегі жәндіктер кәмелетке толмағандар-гормондары тудырды. Молекулалық және жасушалық эндокринология 1, 37-54.
Wogulis, M., Wheelock, CE, Kamita, SG, Hinton, AC, Whetstone, PA, Hammock, BD, Wilson, DK, 2006. Күшті жәндіктердің жетілу ингибиторының құрылымын зерттеу Мандука секстасының кәмелетке толмаған гормонының эстеразасымен байланысқан (, ). Биохимия 45, 4045-4057.
Әрі қарай оқу
- Foucher AL, McIntosh A, Douce G, Wastling J, Tait A, Turner CM (2006). «Трипаносома бруцейіндегі мышьяктың есірткіге төзімділігіне протеомиялық талдау». Протеомика. 6 (9): 2726–32. дои:10.1002 / pmic.200500419. PMID 16526094.
- Mitsui T, Riddiford LM, Bellamy G (1979). «Темекі мүйізтәрізділер (Manduca sexta) эпидермисімен жасөспірім гормонының метаболизмі». Жәндіктер биохимиясы. 9 (6): 637–643. дои:10.1016/0020-1790(79)90103-3.
Әдебиеттер тізімі
- ^ Гилберт және басқалар, 1977 ж
- ^ Нижут және Уильямс, 1974 ж
- ^ Риддифорд, 1976 ж
- ^ Nowock and GILBERT, 1976a; Санбург және басқалар, 1975a; Санбург және басқалар, 1975б
- ^ Ниджхут, 1975 ж
- ^ Хаммак пен Квистад, 1976; Слейд пен Зибитт, 1972 ж
- ^ Винс пен Гилберт, 1977: Искра, 1979 # 1170; Вейрих пен Врен, 1973 ж
- ^ Хаммок және басқалар, 1977; Вейрих пен Врен, 1973; Вейрих пен Рен, 1976 ж
- ^ Хаммок және басқалар, 1975; Новок пен Гилберт, 1976б; Уитмор және басқалар, 1974 ж
- ^ Уитмор, 1972 ж. # 1160; Уитмор және басқалар, 1974 ж
- ^ Крамер, 1978 ж
- ^ Редди және басқалар, 1979 ж
- ^ Джонс және басқалар, 1980
- ^ Sparks and Hammock, 1979 ж
- ^ Sparks және басқалар, 1979 ж
- ^ Джонс және басқалар, 1980
- ^ Sparks және басқалар, 1983 ж
- ^ Хаммок және басқалар, 1977 ж
- ^ Абдель-Аал және басқалар, 1984 ж
- ^ Браун және Уайт, 1995; Де Корт және Грейнжер, 1996; Плапп және басқалар, 1998; Прествич және басқалар, 1996
- ^ Бейкер және басқалар, 1987 ж
- ^ Хвансу және басқалар, 1979 ж
- ^ Спарагана және басқалар, 1985; Спарагана және басқалар, 1984 ж
- ^ Бейкер және басқалар, 1987 ж
- ^ Джонс және басқалар, 1990 ж
- ^ Вогулис және басқалар, 2006