Айна нейрон - Mirror neuron
Бұл мақала болуы керек жаңартылды. (Сәуір 2020) |
| Айна жүйесі | |
|---|---|
| Идентификаторлар | |
| MeSH | D059167 |
| Нейроанатомияның анатомиялық терминдері | |
A айна нейрон Бұл нейрон бұл өрттер хайуан әрекет еткенде де, жануар сол әрекетті басқа орындаған кезде де байқайды.[1][2][3] Осылайша, нейрон бақылаушының өзі әрекет ететіндей, екіншісінің мінез-құлқын «бейнелейді». Мұндай нейрондар адамда тікелей байқалған[дәйексөз қажет ] және примат түрлері,[4] және құстар.[5]
Адамдарда мидың айнадағы нейрондардың белсенділігі сәйкес келеді премоторлы кортекс, қосымша қозғалтқыш аймағы, бастапқы соматосенсорлы қыртыс, және төменгі париетальды қыртыс.[6] Адамдардағы айна жүйесінің қызметі көптеген болжамдардың тақырыбы болып табылады. Құстарда имитациялық-резонанстық мінез-құлық бар екендігі дәлелденді және неврологиялық дәлелдер айна жүйесінің қандай да бір формасының болуын болжайды.[4][7]
Бүгінгі күнге дейін айнадағы нейрондық белсенділіктің когнитивті функцияларды қалай қолдайтынын сипаттайтын бірде-бір жүйке немесе есептеу модельдері ұсынылған жоқ.[8][9][10] Айна нейрондарының тақырыбы қызу пікірталас туғызуда. 2014 жылы, Корольдік қоғамның философиялық операциялары B толығымен айналы нейрондық зерттеулерге арналған арнайы шығарылым шығарды.[11]
Когнитивті неврологиядағы кейбір зерттеушілер және когнитивті психология бұл жүйе қабылдау / іс-қимыл байланысының физиологиялық механизмін қамтамасыз етеді деп есептеңіз (қараңыз жалпы кодтау теориясы ).[3] Олар айнадағы нейрондар басқа адамдардың іс-әрекеттерін түсіну үшін және еліктеу арқылы жаңа дағдыларды үйрену үшін маңызды болуы мүмкін деп тұжырымдайды. Кейбір зерттеушілер айна жүйелері байқалған әрекеттерді имитациялайды, сөйтіп үлес қосуы мүмкін деп болжайды ақыл теориясы дағдылар,[12][13] ал басқалары айна нейрондарын байланыстырады тіл мүмкіндіктер.[14] Марко Якобони (UCLA) сияқты нейробиологтар айнадағы нейрондық жүйелерді адамның миы басқа адамдардың әрекеттері мен ниеттерін түсінуге көмектесіңіз. Якобони және оның әріптестері 2005 жылғы наурызда жарияланған зерттеуде айнадағы нейрондар шай ішіп отырған басқа адамның одан шырын ішуді немесе оны үстелден тазартуды жоспарлап отырғанын біле алады деп хабарлады.[15] Сонымен қатар, Якобони айнадағы нейрондар адамның эмоцияларға қабілеттілігінің жүйке негізі болып табылады деп тұжырымдады эмпатия.[16]
Айна нейрондарының қызметін түсіндіру үшін алға тартылған теорияларға күмәнмен қарайтын ғалымдар бар. Арналған 2013 мақаласында Сымды, Кристиан Джаррет мынаны ескертті:
... айнадағы нейрондар - бұл қызықты, қызықты жаңалық, бірақ бұларды бұқаралық ақпарат құралдарында көргенде, осы жасушалардағы зерттеулердің көпшілігі маймылдарда жүргізілгенін ұмытпаңыз. Айна нейронының әр түрлі түрлері бар екенін ұмытпаңыз. Біз олардың адамдарда бар-жоғын және олардың маймылдардың нұсқаларымен салыстыруын әлі де анықтауға тырысамыз. Бұл жасушалардың функционалдық маңыздылығын түсіну туралы ... алданбаңыз: бұл саяхат енді басталды.[17]
Ашу
1980-1990 жылдары нейрофизиологтар Джакомо Риццолатти, Джузеппе Ди Пеллегрино, Лучано Фадига, Леонардо Фогасси және Vittorio Gallese кезінде Парма университеті орналастырылған электродтар ішінде вентральды премоторлы кортекс туралы макака маймыл оқуға нейрондар қол мен ауыз қуысының әрекеттерін басқаруға мамандандырылған; мысалы, нысанды ұстап алу және оны манипуляциялау. Әр эксперимент кезінде зерттеушілер маймылға тамақ бөліктеріне қол жеткізуге мүмкіндік берді және маймылдың миындағы жалғыз нейрондардан жазып алды, осылайша нейронның белгілі бір қимылдарға реакциясын өлшеді.[18][19] Олардың пайымдауынша, кейбір нейрондар маймыл адамның бір тамақты алып жатқанын көргенде, сондай-ақ маймыл өзі тамақты алып жатқанда, жауап берген. Табиғат, бірақ бәсекелестігі азырақ журналда жарияланғанға дейін «жалпы қызығушылықтың жоқтығы» үшін қабылданбады.[20]
Бірнеше жылдан кейін дәл сол топ тағы бір эмпирикалық мақаланы жариялады, іс-әрекетті танудағы айна-нейрондық жүйенің рөлін талқылап, адамға Брока аймағы болды гомолог маймылдардың вентральды премоторлы кортексінің аймағы.[21]Бұл құжаттарда қолдың іс-әрекетіне жауап беретін айна нейрондарының болуы туралы айтылған болса, Пьер Франческо Феррари мен оның әріптестерінің келесі зерттеуі[22] ауыз қимылдарына және бет қимылдарына жауап беретін айна нейрондарының болуын сипаттады.
Әрі қарайғы тәжірибелер маймылдың төменгі фронтальды және төменгі париетальды кортексіндегі 10% -дық нейрондардың «айна» қасиеттеріне ие екендігін және қолдың орындалған әрекеттері мен бақыланатын әрекеттеріне ұқсас жауаптар беретіндігін растады. 2002 жылы Христиан Кизерс және әріптестердің айтуынша, адамдарда да, маймылдарда да айна жүйесі әрекеттердің дыбысына жауап береді.[3][23][24]
Айнадағы нейрондар туралы есептер кеңінен жарияланды[25] және расталды[26] мидың төменгі фронтальды және төменгі париетальды аймақтарында орналасқан айна нейрондарымен. Жақында функционалды нейро бейнелеу адамдардың ұқсас айналы нейрондық жүйелері бар екенін қатты ұсынады: зерттеушілер әрекет кезінде де, бақылау кезінде де жауап беретін ми аймақтарын анықтады. Бұл ми аймақтары макака маймылында кездесетіні таңқаларлық емес[1] Алайда, функционалды магнитті-резонанстық бейнелеу (fMRI) бүкіл миды бірден зерттей алады және ми аймағының әлдеқайда кең желісі адамдарда айна қасиеттерін бұрын ойлағаннан гөрі көрсетеді деп болжайды. Бұл қосымша бағыттарға мыналар жатады соматосенсорлы қыртыс және бақылаушыға бақыланатын жолмен қозғалу қандай болатынын сездіреді деп ойлайды.[27][28]
Шығу тегі
Көптеген адамдар айна нейрондарының айналығы ең алдымен тұқым қуалайтын генетикалық факторларға байланысты және Mirror нейронын дамытуға генетикалық бейімділік дамыды, өйткені олар әрекетті түсінуді жеңілдетеді деп болжайды.[29] Керісінше, бірқатар теориялық мәліметтер айнадағы нейрондар жай білімді ассоциациялардың, соның ішінде, пайда болуы мүмкін екенін айтады Хеббиан теориясы,[30] The Ассоциативті оқыту Теория,[29] Канализация [31] және Қоныс аудару.[дәйексөз қажет ]
Маймылдарда

Зерттеушілер айна нейрондарын жеке зерттеген алғашқы жануар - бұл макака маймылы. Бұл маймылдарда айна нейрондары төменгі фронтальды гирус (F5 аймағы) және төменгі париетальды лобула.[1]
Айна нейрондары басқа жануарларды түсінуге делдал болады деп саналады мінез-құлық. Мысалы, маймыл қағазды жұлып алған кезде жанатын айнадағы нейрон маймыл адамның қағазды жұлып алғанын көргенде немесе қағаздың жыртып жатқанын естігенде де (визуалды белгілерсіз) өртенетін болады. Бұл қасиеттер зерттеушілерді айнадағы нейрондар «қағазды жырту» сияқты әрекеттердің дерексіз ұғымдарын кодтайды деп сендіруге мәжбүр етті, бұл әрекетті маймыл немесе басқа жануар жасайды.[1]
Макакалардағы айна нейрондарының қызметі белгісіз болып қалады. Ересек макакалар еліктеу арқылы үйренбейтін сияқты. Ферраридің және оның әріптестерінің соңғы тәжірибелері нәресте макакалары адамның бет қимылдарына еліктей алады, дегенмен жаңа туған нәрестелер және уақытша шектеулі терезе кезінде.[32] Егер ол әлі эмпирикалық түрде көрсетілмеген болса да, айнадағы нейрондар осы мінез-құлықты және басқа имитациялық құбылыстарды тудырады деген болжам жасалды.[33] Шынында да, маймылдардың имитациялық мінез-құлықты көрсету деңгейі туралы түсінік шектеулі.[8]
Ересек маймылдарда айнадағы нейрондар маймылға басқа маймылдың не істеп жатқанын түсінуге немесе басқа маймылдың әрекетін тануға мүмкіндік беруі мүмкін.[34]
Адамдарда
Әдетте адам миындағы жалғыз нейрондарды зерттеу мүмкін емес, сондықтан адамдардағы айна нейрондарының көптеген дәлелдері жанама болып табылады. Қолдану арқылы миды бейнелеу тәжірибелері функционалды магнитті-резонанстық бейнелеу (фМРТ) адамның екенін көрсетті төменгі маңдай қыртысы және жоғарғы париетальды лоб адам іс-әрекетті жасаған кезде, сондай-ақ адам іс-әрекетті орындайтын басқа жеке адамды көргенде де белсенді болады. Бұл ми аймағында айна нейрондары бар деген болжам жасалды және олар адамның айна нейрон жүйесі ретінде анықталды.[35] Жақында жүргізілген тәжірибелер көрсеткендей, фМРТ көмегімен сканерленген жалғыз қатысушылардың деңгейінде де көптеген фМРИ воксельдері бар үлкен аумақтар әрекеттерді бақылау кезінде де, орындау кезінде де белсенділіктерін арттырады.[27]
Іс-әрекеттің білімін, пантомималық интерпретацияны және биологиялықты тудыратын зақымдану аймақтарын қарастыратын нейропсихологиялық зерттеулер қозғалысты қабылдау Дефициттер төменгі фронталь гирустың тұтастығы мен осы мінез-құлық арасындағы себепті байланысты көрсетті.[36][37][38] Транскраниальды магниттік ынталандыру зерттеулер мұны да растады.[39][40] Бұл нәтижелер айнадағы нейронмен байланысты аймақтардағы белсенділіктің эпифеноменальды болуы екіталай екендігін көрсетеді.
2010 жылдың сәуірінде жарияланған зерттеуде адамның миында айна қасиеттері бар жалғыз нейрондардың жазбалары туралы баяндалады.[41] Мукамель т.б. (Current Biology, 2010) Рональд Рейган UCLA медициналық орталығында эпилепсияға байланысты емделіп жатқан 21 пациенттің миынан жазылған. Пациенттерге хирургиялық емдеу үшін ұстама ошақтарын анықтау үшін интракраниальды тереңдіктегі электродтар салынды. Электродтың орналасуы тек клиникалық критерийлерге негізделген; зерттеушілер пациенттердің келісімімен сол электродтарды өздерінің зерттеулерін «пигбебка» жасау үшін қолданды. Зерттеушілер жеке адам тапсырманы орындаған кезде де, тапсырманы бақылаған кезде де ең үлкен белсенділік танытатын нейрондардың аз санын тапты. Басқа нейрондардың айнаға қарсы қасиеттері болды, яғни олар қатысушы әрекет жасаған кезде жауап берді, бірақ қатысушы бұл әрекетті көргенде тежелді.
Табылған айна нейрондары қосымша қозғалтқыш аймағында және медиальды уақытша кортексте орналасқан (мидың басқа аймақтары таңдалмаған). Таза практикалық себептерге байланысты бұл аймақтар маймылда айнадағы нейрондар тіркелген аймақтармен бірдей емес: Пармадағы зерттеушілер вентральды премоторлы қабықты және онымен байланысты төменгі париетальды лобты зерттеді, эпилепсия сирек кездесетін екі аймақ және демек, бұл аймақтардағы бір жасушалық жазбалар әдетте адамдарда жасалмайды. Екінші жағынан, ешкім әлі күнге дейін маймылдағы қосымша мотор аймағындағы айна нейрондарын немесе медиальды уақытша лобты іздеген жоқ. Сонымен, бұл адамдар мен маймылдардың әртүрлі жерлерде айналы нейрондары бар дегенді білдірмейді, керісінше оларда маймылда жазылған вентральды премоторлы қабықта және төменгі париетальды лобта айналы нейрондар болуы мүмкін. қозғалтқыш аймақтары және оларда адамда тіркелген медиальды уақытша лоб - әсіресе адамның фМРИ-дің егжей-тегжейлі талдауы осы аймақтардың барлығында айна нейрондарының болуымен үйлесімділікті ұсынады.[27]
Тағы бір зерттеуде адамдарда маймылдардан гөрі айна нейрондары көп болмауы керек, керісінше іс-әрекетті бақылау мен орындауда қолданылатын айна нейрондарының негізгі жиынтығы бар деген болжам жасалды. Алайда, айна нейрондарының ұсынылған басқа функциялары үшін айна жүйесі есту, соматосенсорлық және аффективті компоненттерін жасау кезінде мидың басқа аймақтарын жинау мүмкіндігіне ие болуы мүмкін.[42]
Кеміргіштерде
Бірқатар зерттеулер көрсеткендей, егеуқұйрықтар мен тышқандар басқа кеміргіштерге аяқ соққыларын алған кезде қайғы-қасірет белгілерін көрсетеді.[43] Тобы Христиан Кизерс егеуқұйрықтар ауырсынуды сезінгенде немесе басқалардың ауырғанын көргенде және егеуқұйрықтың алдыңғы цингула қыртысында ауырсыну айна нейрондарының болуын анықтады, яғни жануар ауырсыну кезінде де, басқалардың ауырғанына куә болған кезде де жауап беретін нейрондардан.[44] Цингула қыртысының осы аймағын ажырату егеуқұйрықтардағы эмоционалды жұқпаның төмендеуіне әкелді, сондықтан бақылаушы егеуқұйрықтар тағы бір егеуқұйрықтардың ауырсынуына куә бола отырып, аз стрессті көрсетті.[44] Алдыңғы сингулярлы кортекстің гомологты бөлігі адамдардағы ауырсыну сезіміне байланысты,[45] кеміргіштердегі эмоционалды жұқпалы және адамдардың ауырсынуына эмпатияға қатысатын жүйелер арасындағы гомологияны ұсыну.
Айна нейрондарына қатысты күмән
Ғылыми ортада көптеген адамдар айналы нейрондардың ашылуына байланысты толқуларын білдіргенімен, адамдардағы айналы нейрондардың бар екендігіне де, рөліне де күмәнданған ғалымдар бар. Хикок, Пасколо, Динштейн сияқты ғалымдардың айтуы бойынша, айна нейрондары жасушалардың нақты класын құра ма (бұл басқа функцияларға ие жасушаларда кездесетін кездейсоқ құбылысқа қарағанда) анық емес,[46] және айна әрекеті реакцияның нақты түрі ме, әлде мотор жүйесін жеңілдетудің қарапайым артефактісі ме.[9]
2008 жылы Илан Динштейн және т.б. бастапқы анализдер жасушалардың жеке қасиеттерінің сапалық сипаттамаларына негізделгендіктен және қозғалтқыш аймағындағы қатты айна-селективті нейрондардың аздығын ескермегендіктен, олар сенімді емес деп тұжырымдады.[8] Басқа ғалымдар нейрондық өрттің кешігуін өлшеу реакциялардың стандартты уақыттарымен үйлеспейтін сияқты көрінеді,[46] және F5-тегі қозғалтқыш аймағының үзілуі әрекетті танудың төмендеуіне әкеледі деп ешкім айтпағанын атап өтті.[9] (Бұл аргументтің сыншылары бұл авторлар адамның жүйке-психологиялық және TMS осы салалардың бұзылғандығы туралы есептер шынымен де әрекет тапшылығын тудырады[37][39] қабылдаудың басқа түрлеріне әсер етпей.)[38]
2009 жылы Лингнау және басқалар. эксперимент жүргізді, онда олар алдымен байқалған, содан кейін орындалған қозғалтқыш әрекеттерін алдымен орындалған, содан кейін байқалған қозғалтқыш әрекеттерін салыстырды. Олар екі процестің арасында айна нейрондарының адамда жоқтығын көрсететін айтарлықтай асимметрия бар деген қорытындыға келді. Олар «өте маңызды, біз алдымен орындалған, содан кейін байқалған қозғалтқыш әрекеттері үшін бейімделу белгілерін таппадық. Орындалған және бақыланатын қозғалтқыш актілері үшін кросс-модальді бейімделудің табылмауы осы әрекетті қолдайтын айнаның нейрондық теориясымен сәйкес келмейді. тану және түсіну моторлық модельдеуге негізделген ».[47] Алайда, сол жылы Килнер және т.б. егер мақсатқа бағытталған әрекеттер тітіркендіргіш ретінде қолданылса, IPL де, премоторлы аймақтар да бақылау мен орындау арасындағы айна нейрондары болжайтын қайталанудың басылуын көрсетеді.[48]
2009 жылы Грег Хикок айна нейрондарының іс-әрекетті түсінуге қатысады деген пікіріне қарсы кең дәлел келтірді: «Маймылдар мен адамдардағы нейрондық іс-қимыл теориясының сегіз мәселесі». Ол «бұл жасушалар іс-әрекетті түсінуге негізделеді деген алғашқы гипотеза, сонымен қатар, қызықты және қарапайым ақылға қонымды идея. Алайда кеңінен қабылданғанына қарамастан, бұл ұсыныс ешқашан маймылдарда жеткілікті деңгейде тексерілмеген, ал адамдарда мықты эмпирикалық дәлелдер бар талапқа қарсы физиологиялық және жүйке-психологиялық (қосарланған) диссоциациялардың нысаны ».[9]
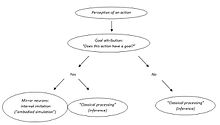
Владимир Косоногов тағы бір қайшылықты көреді. Іс-әрекетті түсінудің айна нейрондық теориясының жақтаушылары айна нейрондары басқалардың іс-әрекеттерінің мақсаттарын кодтайды деп тұжырымдайды, өйткені бақыланатын әрекет мақсатқа бағытталған болса, олар белсендіріледі. Алайда, айнадағы нейрондар бақыланатын іс-әрекет мақсатқа бағытталған кезде ғана іске қосылады (нысанға бағытталған әрекет немесе коммуникативті ым, оның да мақсаты бар). Олар белгілі бір іс-әрекеттің мақсатқа бағытталғандығын қайдан біледі? Белсендірудің қандай кезеңінде олар қозғалыс мақсатын немесе оның жоқтығын анықтайды? Оның пікірінше, айнадағы нейрондық жүйені бақыланатын әрекеттің мақсаты мидың кейбір басқа құрылымдары қойғаннан кейін ғана іске қосуға болады.[49]
Сияқты нейрофилозоферлер Патриция Черчланд айналы нейрондар басқалардың ниеттерін түсінуге жауапты деген теорияға ғылыми және философиялық қарсылықтарын білдірді. 2011 жылғы «Брейнтруст» кітабының 5-тарауында Черчланд айнадағы нейрондардың ниеттерді түсінуге қатысатындығы (бақыланатын әрекеттерді имитациялау арқылы) шешілмеген философиялық мәселелермен көмескіленген болжамдарға негізделгенін көрсетеді. Ол ниеттер жекелеген нейрондарға қарағанда жүйке қызметінің анағұрлым күрделі деңгейінде түсініледі (кодталады) деген дәлелді келтіреді. Черчланд: «Нейрон есептеу жағынан күрделі болғанымен, ол жай нейрон ғана. Бұл интеллектуалды гомункул емес. Егер жүйке торы күрделі нәрсені, мысалы, [қорлау) ниетін білдірсе, ол дұрыс кіріске ие болуы керек және бұл үшін жүйке тізбегіндегі дұрыс орын. « [50]
Жақында Сесилия Хейес (эксперименттік психология профессоры, Оксфорд) айналы нейрондар эволюциялық бейімделуге қарағанда ассоциативті оқытудың қосымша өнімі болып табылады деген теорияны алға тартты. Оның пайымдауынша, адамдардағы айнадағы нейрондар - бұл әрекеттесуді түсіну үшін эволюциялық бейімделу емес, әлеуметтік өзара әрекеттесудің өнімі. Атап айтқанда, Хейс В.С. алға шығарған теорияны жоққа шығарады. Рамачандран айнадағы нейрондар «адам эволюциясындағы үлкен секірістің қозғаушы күші» болды.[10][51]
Даму
Көзді бақылау шараларын қолданатын адамның нәресте деректері айнадағы нейрондық жүйенің 12 айға дейін дамитынын және бұл жүйе адамның сәбилеріне басқа адамдардың іс-әрекеттерін түсінуге көмектесетінін көрсетеді.[52] Айнадағы нейрондардың айна қасиеттерін қалай иемденуіне қатысты маңызды мәселе. Бір-бірімен тығыз байланысты екі модель айна нейрондары арқылы оқытылады деп тұжырымдайды Хеббиан[53] немесе Ассоциативті оқыту[54][55][56] (қараңыз Ассоциативті жүйелілікпен оқыту ). Алайда, егер айна қасиеттеріне ие болу үшін премоторлы нейрондарды іс-әрекет арқылы жаттықтыру қажет болса, жаңа туылған нәрестелер басқа адамның бет қимылдарын қалай имитациялай алатындығы түсініксіз (көрінбейтін әрекеттерге еліктеу) Мельтзофф және Мур. Мүмкіндіктердің бірі - тілдің шығыңқы көрінісі ан босату механизмі жаңа туған нәрестелерде. Мұқият талдау бұл жалғыз қимылға «еліктеу» жаңа туылған нәрестелердің бет-әлпеті туралы барлық дерлік мәліметтерді қамтуы мүмкін екенін көрсетеді.[57]
Мүмкін болатын функциялар
Ниеттерді түсіну
Көптеген зерттеулер айна нейрондарын мақсаттар мен ниеттерді түсінумен байланыстырады. Фогасси және басқалар. (2005)[58] екі резус-макаканың төменгі париетальды лобында (IPL) 41 айна нейрондарының белсенділігін тіркеді. IPL ұзақ уақыт бойы сенсорлық ақпаратты біріктіретін ассоциациялық кортекс ретінде танылды. Маймылдар экспериментаторды алманы ұстап, оны аузына алып келгенін немесе бір затты ұстап, тостағанға салып жатқанын бақылап отырды.
- Жалпы алғанда, 15 айна нейрондары маймыл «жеуге түсіну» қозғалысын бақылаған кезде қатты атқан, бірақ «түсіну-ұстау» жағдайында ешқандай белсенділікті тіркеген жоқ.
- 4 басқа айна нейрондары үшін керісінше болды: олар экспериментатордың соңында алманы тостағанға салғанына жауап берді, бірақ оны жеуге болмайды.
Заттардың манипуляциясы бар кинематикалық күш емес, тек әрекет түрі, нейрондық белсенділік анықталды. Сондай-ақ, маймылға дейін атылған нейрондардың екінші қозғалтқыш әрекетін бастайтын адам нысанын бақылағаны маңызды болды (затты ауызға әкелу немесе кесеге салу). Сондықтан IPL нейрондары «сол әрекетті (түсінуді) әрекетті енгізілген әрекеттің соңғы мақсатына сәйкес басқаша кодтайды». [58] Олар басқа адамның келесі әрекеттерін болжауға және ниетін шығаруға жүйке негізін жасай алады.[58]
Оқытуды жеңілдету
Айна нейрондарының тағы бір мүмкін функциясы - бұл оқытуды жеңілдету. Айна нейрондары іс-әрекеттің нақты көрінісін, яғни бақылаушы әрекет етсе, іске қосылатын бейнені кодтайды. Бұл бақыланатын іс-әрекеттің өзіндік моторлық бағдарламаларын жинау және кейінірек әрекеттерді көбейтуге дайын болу үшін бақыланған әрекетті имитациялық түрде (іште қайталауға) мүмкіндік береді. Бұл жасырын оқыту. Осыған байланысты бақылаушы іс-әрекетті анық (өзінің мінез-құлқында) ептілік пен нәзіктікпен жасайды. Бұл ассоциативті оқыту процестеріне байланысты болады. Синаптикалық байланыс қаншалықты жиі белсендірілсе, соғұрлым ол күшейе түседі.[49]
Эмпатия
Стефани Престон және Франс де Ваал,[59] Жан Дети,[60][61] және Vittorio Gallese[62][63] және Христиан Кизерс[3] айнадағы нейрондық жүйенің қатысатындығын өздері дәлелдеп берді эмпатия. ФМРТ қолданатын көптеген эксперименттер, электроэнцефалография (EEG) және магнетоэнцефалография (MEG) мидың кейбір аймақтары (атап айтқанда, алдыңғы) екенін көрсетті инсула, алдыңғы цингула қыртысы, және төменгі фронтальды кортекс) адамдар эмоцияны сезінгенде (жиіркеніш, бақыт, ауырсыну және т.б.) және эмоцияны бастан өткерген басқа адамды көргенде белсенді болады.[64][65][66][67][68][69][70]Дэвид Фридберг және Vittorio Gallese айналы нейрондық жүйенің бұл қызметі өте маңызды деген идеяны алға тартты эстетикалық тәжірибе.[71] Алайда, мидың бұл аймақтары қолдың іс-әрекетін бейнелейтінімен бірдей емес, ал эмоционалды күйге немесе эмпатияға арналған айна нейрондары әлі маймылдарда сипатталмаған.
Христиан Кизерс Әлеуметтік ми зертханасы және әріптестер өзін-өзі есеп беру сауалнамаларына сәйкес эмпатияға ие адамдар қолмен жұмыс жасау үшін айна жүйесінде де белсенділігі жоғары болатындығын көрсетті[72] және эмоцияларға арналған айна жүйесі,[69] Кейбір зерттеушілер адамның айна жүйесі іс-әрекетті бақылауға пассивті жауап бермейді, бірақ бақылаушының ақыл-ойының әсерінен болады деп байқаған.[73] Зерттеушілер пациенттерді күтуге эмпатикалық қатысу кезінде айна нейрондарының байланысын байқады.[74]
Егеуқұйрықтарға жүргізілген зерттеулер бұл алдыңғы цингула қыртысы құрамында ауырсынуға арналған айна нейрондары бар, яғни ауырсыну сезімі кезінде де, басқалардың ауырғанын көргенде де жауап беретін нейрондар,[44] және осы аймақтың тежелуі төмендеуге әкеледі эмоционалды жұқпа егеуқұйрықтарда[44] және тышқандар,[43] және басқаларға зиян келтіруден бас тартуды азайтты.[75] Бұл ауырсыну айна нейрондарының арасындағы байланысты дәлелді дәлелдермен қамтамасыз етеді эмоционалды жұқпа және прокурорлық мінез-құлық, эмпатиямен байланысты екі құбылыс, кеміргіштерде. Мидың гомологиялық аймағындағы белсенділігі адамдардағы эмпатияның жеке өзгергіштігімен байланысты[45] ұқсас механизм сүтқоректілерде ойнауы мүмкін деп болжайды.
Адамның өзін-өзі тануы
Рамачандран айнадағы нейрондар адамның өзін-өзі тануының неврологиялық негізін қамтамасыз етуі мүмкін деген болжам жасады.[76] 2009 жылы Edge Foundation үшін жазған очеркінде Рамачандран оның теориясын келесідей түсіндірді: «... Мен сондай-ақ, бұл нейрондар басқа адамдардың мінез-құлқын модельдеуге көмектесіп қана қоймай, оларды« ішке »айналдыруы мүмкін деп ойладым. өзіңіздің екінші ретті ұсыныстарыңызды немесе мета-өкілдіктеріңізді жасаңыз меншікті ертерек ми процестері. Бұл интроспекцияның және өзін-өзі танудың және басқа да хабардарлықтың өзара байланысының жүйке негізі болуы мүмкін. Бұл жерде бірінші кезекте эволюцияға байланысты тауық немесе жұмыртқа туралы сұрақ туындайтыны анық, бірақ ... Ең бастысы, екеуі бірігіп дамыды, бір-бірін өзара байытып, қазіргі заманғы адамдарға тән өзіндік жетілген көріністі құрды ». [77]
Тіл
Адамдарда функционалды МРТ зерттеулерінде мидың гипотезаланған тілдік аймақтарының бірі - Брока аймағына жақын маңдайдың төменгі қабығындағы маймылдардың айна нейрондық жүйесіне гомологиялық бағыттар табылғандығы туралы хабарлады. Бұл адам тілінің айна нейрондарында жүзеге асырылатын ым-ишара орындау / түсіну жүйесінен дамығандығы туралы ұсыныстарға әкелді. Айна нейрондарының іс-әрекетті түсіну, еліктеу-үйрену және басқа адамдардың мінез-құлқын модельдеу механизмін қамтамасыз етуге мүмкіндігі бар деп айтылды.[78] Бұл гипотезаны кейбіреулер қолдайды цитоархитектоникалық маймылдардың F5 премоторлы аймағы мен адамның Брока аймағы арасындағы гомология.[79] Ставкалары лексика қабілетіне кеңейту сілтемесі балалар сөздерді емес дыбыстық бейнелеуді және сол сияқты жаңа сөздердің айтылу ерекшеліктерін алуды. Мұндай сөйлеуді қайталау автоматты түрде, жылдам жүреді[80] және миға бөлек сөйлеуді қабылдау.[81][82] Сонымен қатар, мұндай вокалды еліктеу, мысалы, түсінбей пайда болуы мүмкін сөйлеу көлеңкесі[83] және эхолалия.[84]
Бұл байланыстың тағы бір дәлелі жақында жүргізілген зерттеуден туындайды, онда екі қатысушының ми белсенділігі фМРТ көмегімен өлшенді, олар бір-бірімен ойын арқылы қол қимылдарын қолданып сөйлескен кезде акциялар - кейбіреулер ұсынған модальдылық адам тілінің эволюциялық ізашары болуы мүмкін. Деректерді қолдану арқылы талдау Грейнджердің себебі бақылаушының айна-нейрондық жүйесі шынымен де сөзбен байланысты мотор ұғымы айна жүйесінің көмегімен бір мидан екіншісіне беріледі деген идеяны қолдай отырып, жөнелтушінің қозғалтқыш жүйесіндегі белсенділікті бейнелейтіндігін анықтады.[85]
Айнадағы нейрондық жүйе кез-келген рөл атқаруға жеткіліксіз сияқты синтаксис иерархиялық рекурсивті құрылымда жүзеге асырылатын адам тілдерінің осы анықтайтын қасиеті фонемалардың сызықтық тізбектеріне тегістелгенін ескере отырып, рекурсивті құрылымды сенсорлық анықтауға қол жетімсіз етеді.[86]
Автоматты имитация
Бұл термин көбінесе дене қимылын байқаған жеке адам дене қимылын байқамай орындайды немесе дене қимылын өзгерту тәсілін өзгертетін жағдайларға қатысты қолданылады. Автоматты еліктеу сәйкес жауаптардың айқын орындалуын сирек қамтиды. Оның орнына әсерлер әдетте дәлдіктен емес, үйлесімді және үйлесімсіз сынақтар арасындағы айырмашылықтардан тұрады, зерттеу нәтижелері бойынша имитацияның жасырын формасы болып табылатын автоматты имитацияның кеңістіктегі үйлесімділіктен айырмашылығы бар. Сондай-ақ, бұл автоматты имитация зейінді процестермен енгізу модуляциясына және тежеуші процестермен шығумен модуляцияға ұшырағанымен, оны тікелей қасақана процестермен өзгерту мүмкін емес, ұзақ мерзімді сенсомоторлы ассоциациялар арқылы жүзеге асырылатындығын көрсетеді. Көптеген зерттеушілер автоматты имитацияны айнадағы нейрон жүйесі жасайды деп санайды.[87] Сонымен қатар, адамдар басқа әрекеттер туралы сөйлемдерді тыңдағанда, біздің постуральды бақылауымыздың нашарлайтындығын көрсететін деректер бар. Мысалы, егер дене бітімін сақтау міндеті болса, адамдар келесі сөйлемдерді тыңдағанда мұны нашарлатады: «Мен тұрамын, тәпішкені киемін, жуынатын бөлмеге барамын». Бұл құбылыс әрекетті қабылдау кезінде адамда бір әрекетті жасағандай (мылжың нейрондар жүйесі) моторлы кортекстің активтенуі болатындығына байланысты болуы мүмкін.[88]
Қозғалтқыштың мимикасы
Автоматты имитациядан айырмашылығы, моторлық имитация (1) натуралистік әлеуметтік жағдайларда және (2) сынақтардағы жылдамдық және / немесе дәлдік өлшемдерінен гөрі сеанс ішіндегі әрекет жиілігі өлшемдері арқылы байқалады.[89]
Қозғалтқыш мимикасы мен автоматты имитацияны зерттеудің интеграциясы бұл құбылыстардың бірдей психологиялық және жүйке процестеріне тәуелді екендігінің дәлелді белгілерін анықтай алады. Алдын ала дәлелдемелер әлеуметтік примингтің моторлық мимикаға ұқсас әсері бар екенін көрсететін зерттеулерден алынған.[90][91]
Осыған қарамастан, автоматты еліктеу, айна эффектілері және моторлық имитация арасындағы ұқсастықтар кейбір зерттеушілерге автоматты имитацияны айналы нейрондық жүйенің көмегімен жүзеге асырады және бұл натуралистік әлеуметтік контексте байқалатын моторлы мимиканың қатаң бақыланатын зертханалық эквиваленті болып табылады деген ұсыныс жасады. Егер рас болса, онда автоматты имитация айнадағы нейрондық жүйенің когнитивтік қызметке қаншалықты ықпал ететіндігін және моторлы мимиканың просоциалдық көзқарастар мен мінез-құлықты қалай дамытатынын зерттеу құралы ретінде қолданыла алады.[92][93]
Адамдардағы имитациялық зерттеулердің мета-анализі еліктеу кезінде айна нейронының қатысуы ықтимал болатын айна жүйесінің активтенуі туралы жеткілікті дәлелдер бар екенін көрсетеді, дегенмен жарияланған сингулярлық нейрондардың белсенділігі туралы ешқандай жарияланымдар жоқ. Дегенмен, бұл моторды имитациялау үшін жеткіліксіз болуы мүмкін. Зерттеулер көрсеткендей, имитация кезінде классикалық айна жүйесінен тыс орналасқан фронталь және париетальды лобтардың аймақтары бірдей белсенді болады. Бұл имитациялық мінез-құлық үшін айна жүйесімен бірге басқа аймақтардың шешуші маңызы бар екенін көрсетеді.[6]
Аутизм
Сондай-ақ, айнадағы нейрондық жүйенің проблемалары когнитивті бұзылулардың негізінде болуы мүмкін деп ұсынылды, әсіресе аутизм.[94][95] Алайда, айнаның нейрондық дисфункциясы мен аутизм арасындағы байланыс болжамды болып табылады және айна нейрондарының аутизмнің көптеген маңызды сипаттамаларымен байланысын көрсету қажет.[8]
Кейбір зерттеушілер айна нейрондарының жетіспеушілігі мен арасында байланыс бар деп мәлімдейді аутизм. Қозғалтқыш аймағындағы ЭЭГ жазбалары біреу басқа адамның қимылын қараған кезде басылады, бұл айнадағы нейрондық жүйеге қатысты сигнал болуы мүмкін. Аутизмі бар балаларда бұл басу аз болды.[94] Бұл тұжырымдарды бірнеше топ қайталағанымен,[96][97] басқа зерттеулер аутизмде дисфункционалды айна нейрондық жүйесінің дәлелін таппады.[8] 2008 жылы Оберман және басқалар. қарама-қайшы ЭЭГ дәлелдерін ұсынған зерттеу жұмысын жариялады. Оберман мен Рамачандран таныс емес тітіркендіргіштер үшін емес, әдеттегі му-супрессияны тапты, сондықтан оларды ASD (аутизм спектрі бұзылуы) бар балалардың айна нейрондық жүйесі функционалды, бірақ типтік балаларға қарағанда сезімталдығы төмен деген қорытындыға келді.[98] Му толқынды басу эксперименттері ұсынған қарама-қайшы дәлелдерге сүйене отырып, Патриция Черчланд му толқынды басу нәтижелері айна нейрондық жүйелерінің жұмысын өлшеу үшін жарамды индекс ретінде қолданыла алмайтындығын ескертті.[99] Соңғы зерттеулер айнадағы нейрондардың аутизмде рөл атқармайтынын көрсетті:
... аутизмдегі айна жүйесінің негізгі тапшылығына нақты дәлелдер жоқ. Мінез-құлықты зерттеу көрсеткендей, аутизммен ауыратын адамдар іс-әрекеттің мақсаттарын жақсы түсінеді. Сонымен қатар, екі тәуелсіз нейровизорлық зерттеулер айна жүйесінің париетальды компоненті әдетте аутизммен ауыратын адамдарда жұмыс істейтіндігін хабарлады.[100]
Аутизмді емес ересектермен салыстырғанда аутизм спектрі бұзылған ересектердегі айнадағы нейронмен байланысты ми аймағында кейбір анатомиялық айырмашылықтар табылды. Бұл кортикальды аймақтардың барлығы жұқа және жұқару дәрежесі аутизм симптомдарының ауырлығымен байланысты болды, корреляция бұл ми аймақтарымен шектелді.[101] Осы нәтижелерге сүйене отырып, кейбір зерттеушілер аутизм айнадағы нейрондық жүйенің бұзылуынан туындайды, бұл әлеуметтік дағдыларды, еліктеуді, эмпатияны және ақыл-ой теориясының бұзылуына әкеледі деп мәлімдейді.[ДДСҰ? ]
Көптеген зерттеушілер аутизмнің «сынған айналары» теориясының шамадан тыс қарапайым екендігін және тек айналы нейрондар аутизмі бар адамдарда кездесетін айырмашылықтарды түсіндіре алмайды деп атап көрсетті. Біріншіден, жоғарыда айтылғандай, бұл зерттеулердің ешқайсысы айна нейрондарының белсенділігі болып табылмады - басқаша айтқанда фМРА белсенділігі немесе ЭЭГ ырғағының басылуы айна нейрондарын анық индекстемейді. Динштейн және оның әріптестері аутизммен ауыратын адамдарда фМРИ қолданып, нейронның қалыпты айна белсенділігін анықтады.[102] Аутизмі бар адамдарда ниетті түсінудің, әрекетті түсінудің және қозғалысты биологиялық қабылдаудың тапшылығы (айна нейрондарының негізгі функциялары) әрдайым кездесе бермейді,[103][104] немесе тапсырмаға тәуелді.[105][106] Бүгінгі күні өте аз адамдар айна жүйесіндегі аутизмнің негізінде ештеңе жоқ деп айтуға болады деп санайды. Мұның орнына «қосымша зерттеулер жүргізу керек, ал бұқаралық ақпарат құралдарына хабарласқанда көбірек сақтық қажет». [107]
2010 жылдан бастап зерттеу[108] аутист адамдар айнадағы нейрондық дисфункцияны көрсетпейді деген тұжырымға келді, дегенмен сынаманың кіші мөлшері бұл нәтижелерді жалпылауға мүмкіндік береді.
Ақыл-ой теориясы
Жылы Ақыл-ой философиясы, айнадағы нейрондар негізгі шақыруға айналды модельдеу теоретиктері туралы «ақыл теориясы. «» Ақыл-ой теориясы «біздің басқа адамның психикалық жағдайын (яғни сенім мен тілектерді) тәжірибе немесе олардың мінез-құлқынан шығару қабілетімізді білдіреді.
Біздің ақыл теориямызды есепке алуға тырысатын бірнеше бәсекелес модельдер бар; айналы нейрондарға қатысты ең көрнекті - имитациялық теория. Имитациялық теорияға сәйкес ақыл-ой теориясы қол жетімді, өйткені біз бейсаналық біз бақылап отырған адамға жанашырлықпен қарап, сәйкесінше айырмашылықтарды ескере отырып, біз осы сценарийге не қалайтынымызды және оған сенетінімізді елестетіңіз.[109][110] Mirror neurons have been interpreted as the mechanism by which we simulate others in order to better understand them, and therefore their discovery has been taken by some as a validation of simulation theory (which appeared a decade before the discovery of mirror neurons).[62] More recently, Theory of Mind and Simulation have been seen as complementary systems, with different developmental time courses.[111][112][113]
At the neuronal-level, in a 2015 study by Keren Haroush and Ziv Williams using jointly interacting primates performing an iterated prisoner's dilemma game, the authors identified neurons in the алдыңғы цингула қыртысы that selectively predicted an opponent's yet unknown decisions or covert state of mind. These "other-predictive neurons" differentiated between self and other decisions and were uniquely sensitive to social context, but they did not encode the opponent's observed actions or receipt of reward. These cingulate cells may therefore importantly complement the function of mirror neurons by providing additional information about other social agents that is not immediately observable or known.[114]
Sex-Based Differences
A series of recent studies conducted by Yawei Cheng, using a variety of neurophysiological measures, including MEG,[115] жұлын рефлексті қозғыштығы,[116] electroencephalography,[117][118] have documented the presence of a gender difference in the human mirror neuron system, with female participants exhibiting stronger motor resonance than male participants.
In another study, sex-based differences among mirror neuron mechanisms was reinforced in that the data showed enhanced empathetic ability in females relative to males. During an emotional social interaction, females showed a greater ability in emotional perspective taking than did males when interacting with another person face-to-face. However, in the study, data showed that when it came to recognizing the emotions of others, all participants' abilities were very similar and there was no key difference between the male and female subjects.[119]
Sleep paralysis / Ghostly Bedroom Intruders
Baland Jalal and Рамачандран have hypothesized that the mirror neuron system is important in giving rise to the intruder hallucination and out-of-body experiences during ұйқының салдануы.[120] According to this theory, sleep paralysis leads to disinhibition of the mirror neuron system, paving the way for hallucinations of human-like shadowy beings. The deafferentation of sensory information during sleep paralysis is proposed as the mechanism for such mirror neuron disinhibition.[120] The authors suggest that their hypothesis on the role of the mirror neuron system could be tested:
"These ideas could be explored using neuroimaging, to examine the selective activation of brain regions associated with mirror neuron activity, when the individual is hallucinating an intruder or having an out-of-body experience during sleep paralysis ."[120]
Mirror neuron function, psychosis, and empathy in schizophrenia
Recent research, which measured mu-wave suppression, suggests that mirror neuron activity is positively correlated with psychotic symptoms (i.e., greater mu suppression/mirror neuron activity was highest among subjects with the greater severity of psychotic symptoms). Researchers concluded that "higher mirror neuron activity may be the underpinning of schizophrenia sensory gating deficits and may contribute to sensory misattributions particularly in response to socially relevant stimuli, and be a putative mechanism for delusions and hallucinations."[121]
Сондай-ақ қараңыз
Әдебиеттер тізімі
- ^ а б c г. Риццолатти, Джакомо; Крайгеро, Лайла (2004). «Айна-нейрондық жүйе» (PDF). Неврологияның жылдық шолуы. 27 (1): 169–192. дои:10.1146 / annurev.neuro.27.070203.144230. PMID 15217330.
- ^ Keysers, Christian (2010). «Айна нейрондары» (PDF). Қазіргі биология. 19 (21): R971–973. дои:10.1016 / j.cub.2009.08.026. PMID 19922849. S2CID 12668046. Архивтелген түпнұсқа (PDF) 2013-01-19.
- ^ а б c г. Keysers, Christian (2011-06-23). The Empathic Brain. Kindle.
- ^ а б Риццолатти, Джакомо; Fadiga, Luciano (1999). "Resonance Behaviors and Mirror Neurons". Italiennes de Biologie. 137 (2–3): 85–100. PMID 10349488.
- ^ Prather, J. F.; Петерс, С .; Nowicki, S.; Mooney, R. (2008). "Precise auditory–vocal mirroring in neurons for learned vocal communication". Табиғат. 451 (7176): 305–310. Бибкод:2008Natur.451..305P. дои:10.1038/nature06492. PMID 18202651. S2CID 4344150.
- ^ а б Molenberghs P, Cunnington R, Mattingley JB (July 2009). "Is the mirror neuron system involved in imitation? A short review and meta-analysis". Неврология және биобевиоралдық шолулар. 33 (7): 975–80. дои:10.1016/j.neubiorev.2009.03.010. PMID 19580913. S2CID 25620637.
- ^ Akins, Chana; Klein, Edward (2002). "Imitative Learning in Japanese Quail using Bidirectional Control Procedure". Жануарларды оқыту және мінез-құлық. 30 (3): 275–281. дои:10.3758/bf03192836. PMID 12391793.
- ^ а б c г. e Динштейн I, Томас С, Бехрман М, Хигер DJ (2008). «Табиғатқа дейінгі айна». Curr Biol. 18 (1): R13-8. дои:10.1016 / j.cub.2007.11.004. PMC 2517574. PMID 18177704.
- ^ а б c г. Hickok G (2009). "Eight problems for the mirror neuron theory of action understanding in monkeys and humans". J Cogn Neurosci. 21 (7): 1229–1243. дои:10.1162/jocn.2009.21189. PMC 2773693. PMID 19199415.
- ^ а б Heyes, Cecilia (2009). "Where do mirror neurons come from?" (PDF). Неврология және биобевиоралдық шолулар. Архивтелген түпнұсқа (PDF) 2012-04-26. Алынған 2011-12-20.
- ^ Ferrari, PF; Rizzolatti, G (2014). "Mirror neuron research: the past and the future". Philos Trans R Soc Lond B Biol Sci. 369 (1644): 20130169. дои:10.1098/rstb.2013.0169. PMC 4006175. PMID 24778369.
- ^ Кисерс, христиан; Gazzola, Valeria (2006). «Миды зерттеудегі прогресс» (PDF). Bcn-nic.nl. Архивтелген түпнұсқа (PDF) 2007-06-30.
- ^ Майкл Арбиб, Айна жүйесінің гипотезасы. Тілді ақыл-ой теориясымен байланыстыру, 2005, retrieved 2006-02-17 Мұрағатталды 2009 жылғы 29 наурыз, сағ Wayback Machine
- ^ Терет, Уго; Паскаль-Леоне, Альваро (2002). "Language Acquisition: Do as You Hear". Қазіргі биология. 12 (21): R736-7. дои:10.1016 / S0960-9822 (02) 01251-4. PMID 12419204.
- ^ Iacoboni, Marco (February 22, 2005). "Grasping the Intentions of Others with One's Own Mirror Neuron System". PLOS биологиясы. 3 (3): e79. дои:10.1371 / journal.pbio.0030079. PMC 1044835. PMID 15736981.
- ^ Blakeslee, Sandra (January 10, 2006). «Ақылға салатын жасушалар». New York Times | Ғылым.
- ^ Джарретт, Кристиан, неврологиядағы айна нейрондарындағы ең гипотеза тұжырымдамаға тыныш қарау, сымды, 12.13.13,[1]
- ^ Di Pellegrino, G.; Фадига, Л .; Фогасси, Л .; Галлез, V .; Rizzolatti, G (1992). «Қозғалтқыш оқиғаларын түсіну: нейрофизиологиялық зерттеу». Миды эксперименттік зерттеу. 91 (1): 176–180. дои:10.1007 / bf00230027. PMID 1301372. S2CID 206772150.
- ^ Риццолатти, Джакомо; Фадига, Лучано; Галлез, Витторио; Fogassi, Leonardo (1996). "Premotor cortex and the recognition of motor actions". Миды когнитивті зерттеу. 3 (2): 131–141. дои:10.1016/0926-6410(95)00038-0. PMID 8713554.
- ^ Rizzolatti G, Fabbri-Destro M (January 2010). "Mirror neurons: from discovery to autism". Миды эксперименттік зерттеу. 200 (3–4): 223–37. дои:10.1007 / s00221-009-2002-3. PMID 19760408. S2CID 3342808.
- ^ Галлез, V .; Фадига, Л .; Фогасси, Л .; Риццолатти, Джакомо (1996). «Қабық алдындағы кортекстегі әрекеттерді тану». Ми. 119 (2): 593–609. дои:10.1093 / ми / 119.2.593. PMID 8800951.
- ^ Ferrari; т.б. (2003). "Mirror neurons responding to the observation of ingestive and communicative mouth actions in the ventral premotor cortex". European Journal of Neuroscience. 17 (8): 1703–1714. дои:10.1046 / j.1460-9568.2003.02601.x. PMID 12752388. S2CID 1915143.
- ^ Kohler; т.б. (2002). «Ғылым» (PDF). Bcn-nic.nl. Архивтелген түпнұсқа (PDF) 2007-06-30.
- ^ Gazzola; т.б. (2006). "Current Biology" (PDF). Bcn-nic.nl. Архивтелген түпнұсқа (PDF) 2007-06-30.
- ^ Галлез, Витторио; Фадига, Лучано; Fogassi, Leonardo; Риццолатти, Джакомо (1996). «Қабық алдындағы кортекстегі әрекеттерді тану». Ми. 119 (2): 593–609. дои:10.1093 / ми / 119.2.593. PMID 8800951.
- ^ Fogassi et al., Parietal Lobe: From Action Organization to Intention Understanding, Science, 2005
- ^ а б c Газзола, V .; Keysers, C. (2009). "The observation and execution of actions share motor and somatosensory voxels in all tested subjects: single-subject analyses of unsmoothed fMRI data". Cereb Cortex. 19 (6): 1239–1255. дои:10.1093/cercor/bhn181. PMC 2677653. PMID 19020203.
- ^ Кисерс, христиан; Kaas, John; Gazzola, Valeria (2010). "Somatosensation in Social Cognition" (PDF). Табиғи шолулар неврология. 11 (6): 417–28. дои:10.1038/nrn2833. PMID 20445542. S2CID 12221575. Архивтелген түпнұсқа (PDF) 2010-08-16.
- ^ а б Cook R, Bird G, Catmur C, Press C, Heyes C (2014). "Mirror neurons: from origin to function". Behav Brain Sci. 37 (2): 177–92. дои:10.1017/S0140525X13000903. PMID 24775147.CS1 maint: бірнеше есімдер: авторлар тізімі (сілтеме)
- ^ Keysers C, Gazzola V (2014). "Hebbian learning and predictive mirror neurons for actions, sensations and emotions". Philos Trans R Soc Lond B Biol Sci. 369 (1644): 20130175. дои:10.1098/rstb.2013.0175. PMC 4006178. PMID 24778372.
- ^ Del Giudice M, Manera V, Keysers C (2009). "Programmed to learn? The ontogeny of mirror neurons". Dev Sci. 12 (2): 350–63. дои:10.1111/j.1467-7687.2008.00783.x. hdl:2318/133096. PMID 19143807.CS1 maint: бірнеше есімдер: авторлар тізімі (сілтеме)
- ^ Феррари, П.Ф .; Visalberghi, E.; Паукнер, А .; Фогасси, Л .; Ruggiero, A.; Suomi, SJ; т.б. (2006). "Neonatal Imitation in Rhesus Macaques". PLOS биологиясы. 4 (9): e302. дои:10.1371/journal.pbio.0040302. PMC 1560174. PMID 16953662.
- ^ Ferrari P.F., Bonini L., Fogassi L. (2009) From monkey mirror neurons to mirror-related behaviours: possible direct and indirect pathways. Philosophical Transactions of the Royal Society, B. .364, 2311-2323
- ^ Риццолатти, Джакомо; Arbib, Michael A. (1998). "Language within our grasp". Неврология ғылымдарының тенденциялары. 21 (5): 188–194. дои:10.1016/S0166-2236(98)01260-0. hdl:11858/00-001M-0000-002C-4B59-6. PMID 9610880. S2CID 679023.
- ^ Якобони, Марко; Woods, Roger P.; Brass, Marcel; Bekkering, Harold; Маззиотта, Джон С .; Rizzolatti, Giacomo (1999). "Cortical Mechanisms of Human Imitation" (PDF). Ғылым. 286 (5449): 2526–2528. дои:10.1126 / ғылым.286.5449.2526. PMID 10617472.
- ^ Saygin AP, Wilson SM, Dronkers NF, Bates E (2004). "Action comprehension in aphasia: linguistic and non-linguistic deficits and their lesion correlates". Нейропсихология. 42 (13): 1788–804. CiteSeerX 10.1.1.544.9071. дои:10.1016/j.neuropsychologia.2004.04.016. PMID 15351628. S2CID 11622224.
- ^ а б Транель, Д .; Kemmerer, D.; Adolphs, R.; Дамасио, Х .; Damasio, A. R. (2003-05-01). "Neural correlates of conceptual knowledge for actions". Когнитивті нейропсихология. 20 (3): 409–432. дои:10.1080/02643290244000248. PMID 20957578. S2CID 16131.
- ^ а б Saygin, A. P. (2007). "Superior temporal and premotor brain areas necessary for biological motion perception". Brain : A Journal of Neurology. 130 (Pt 9): 2452–2461. дои:10.1093/brain/awm162. PMID 17660183.
- ^ а б Pobric, G.; De c. Hamilton, A. F. D. C. (2006-03-07). "Action understanding requires the left inferior frontal cortex". Қазіргі биология. 16 (5): 524–529. дои:10.1016/j.cub.2006.01.033. PMID 16527749.
- ^ Candidi M; Urgesi C; Ionta S; Aglioti SM (2008). "Virtual lesion of ventral premotor cortex impairs visual perception of biomechanically possible but not impossible actions". Әлеуметтік неврология. 3 (3–4): 388–400. дои:10.1080/17470910701676269. PMID 18979387. S2CID 37390465.
- ^ Кисерс, христиан; Gazzola, Valeria (2010). "Social Neuroscience: Mirror Neurons recorded in Humans" (PDF). Қазіргі биология. 20 (8): R353–354. дои:10.1016/j.cub.2010.03.013. hdl:20.500.11755/351f0172-b06b-41de-a013-852c64e197fa. PMID 21749952. S2CID 3609747. Архивтелген түпнұсқа (PDF) 2010-08-16.
- ^ Molenberghs P, Cunnington R, Mattingley JB (January 2012). "Brain regions with mirror properties: a meta-analysis of 125 human fMRI studies" (PDF). Неврология және биобевиоралдық шолулар. 36 (1): 341–9. дои:10.1016/j.neubiorev.2011.07.004. PMID 21782846. S2CID 37871374.
- ^ а б Keum S, Shin HS (2019). "Neural Basis of Observational Fear Learning: A Potential Model of Affective Empathy". Нейрон. 104 (1): 78–86. дои:10.1016/j.neuron.2019.09.013. PMID 31600517. S2CID 203932127.
- ^ а б c г. Carrillo M, Han Y, Migliorati F, Liu M, Gazzola V, Keysers C (2019). "Emotional Mirror Neurons in the Rat's Anterior Cingulate Cortex". Curr Biol. 29 (8): 1301–1312.e6. дои:10.1016/j.cub.2019.03.024. PMC 6488290. PMID 30982647.CS1 maint: бірнеше есімдер: авторлар тізімі (сілтеме)
- ^ а б Lamm C, Decety J, Singer T (2011). "Meta-analytic evidence for common and distinct neural networks associated with directly experienced pain and empathy for pain". NeuroImage. 54 (3): 2492–502. дои:10.1016/j.neuroimage.2010.10.014. PMID 20946964. S2CID 6021487.CS1 maint: бірнеше есімдер: авторлар тізімі (сілтеме)
- ^ а б Pascolo PB, Ragogna R, Rossi R (2009). "The Mirror-Neuron System Paradigm and its consistency". Жүру және қалып. 30 (Suppl. 1): 65. дои:10.1016/j.gaitpost.2009.07.064.
- ^ Lingnau, A.; Gesierich, B.; Caramazza, A. (February 28, 2009). "Asymmetric fMRI adaptation reveals no evidence for mirror neurons in humans". Ұлттық ғылым академиясының материалдары. 106 (24): 9925–9930. Бибкод:2009PNAS..106.9925L. дои:10.1073/pnas.0902262106. PMC 2701024. PMID 19497880.
- ^ Kilner, JM.; Neal, A. (August 12, 2009). "Evidence of mirror neurons in human inferior frontal gyrus". J Neurosci. 29 (32): 10153–10159. дои:10.1523/jneurosci.2668-09.2009. PMC 2788150. PMID 19675249.
- ^ а б Kosonogov, V. (2012). "Why the Mirror Neurons Cannot Support Action Understanding". Нейрофизиология. 44 (6): 499–502. дои:10.1007/s11062-012-9327-4. ISSN 0090-2977. S2CID 6315381.
- ^ Churchland, Patricia, Braintrust (2011), Chapter 6, page 142
- ^ Ramachandran, VS (2000). "Mirror neurons and imitation learning as the driving force behind "the great leap forward" in human evolution". Жиек. Алынған 13 сәуір 2013.
- ^ Falck-Ytter T, Gredebäck G, von Hofsten C (2006). "Infants predict other people's action goals" (PDF). Nature Neuroscience 9. Psyk.uu.se. Архивтелген түпнұсқа (PDF) 2009-03-25.
- ^ Keysers & Perrett, Trends in Cognitive Sciences 8 (2004)
- ^ Heyes C. M. (2001). "Causes and consequences of imitation". Когнитивті ғылымдардың тенденциялары. 5 (6): 253–261. дои:10.1016/S1364-6613(00)01661-2. PMID 11390296. S2CID 15602731.
- ^ Brass, M., & Heyes, C. Когнитивті ғылымдардың тенденциялары 9 (2005)
- ^ Heyes C. M. (2010). "Where do mirror neurons come from?". Неврология және биобевиоралдық шолулар. 34 (4): 575–583. дои:10.1016/j.neubiorev.2009.11.007. PMID 19914284. S2CID 2578537.
- ^ Anisfeld M (1996). "Only tongue protruding modeling is matched by neonates". Даму шолу. 16 (2): 149–161. дои:10.1006/drev.1996.0006.
- ^ а б c Fogassi L, Ferrari PF, Gesierich B, Rozzi S, Chersi F, Rizzolatti G (April 2005). "Parietal lobe: from action organization to intention understanding". Ғылым. 308 (5722): 662–7. Бибкод:2005Sci...308..662F. дои:10.1126/science.1106138. PMID 15860620. S2CID 5720234.
- ^ Preston, S. D.; де Ваал, Ф.Б.М. (2002). «Эмпатия: оның түпкілікті және жақын негіздері». Мінез-құлық және ми туралы ғылымдар. 25 (1): 1–72. CiteSeerX 10.1.1.554.2794. дои:10.1017 / s0140525x02000018. PMID 12625087.
- ^ Decety, J. (2002). Naturaliser l’empathie [Empathy naturalized]. L’Encéphale, 28, 9-20.
- ^ Decety J, Jackson PL (2004). «Адам эмпатиясының функционалдық архитектурасы». Неврология туралы мінез-құлық және когнитивті шолулар. 3 (2): 71–100. дои:10.1177/1534582304267187. PMID 15537986.
- ^ а б Gallese V, Goldman A (December 1998). «Айна нейрондары және ойды оқудың имитациялық теориясы». Когнитивті ғылымдардың тенденциялары. 2 (12): 493–501. дои:10.1016 / s1364-6613 (98) 01262-5. PMID 21227300. S2CID 10108122.
- ^ Gallese V (2001). "The "Shared Manifold" hypothesis: from mirror neurons to empathy". Сана туралы зерттеулер журналы. 8: 33–50.
- ^ Botvinick M, Jha AP, Bylsma LM, Fabian SA, Solomon PE, Prkachin KM (2005). "Viewing facial expressions of pain engages cortical areas involved in the direct experience of pain". NeuroImage. 25 (1): 312–319. дои:10.1016/j.neuroimage.2004.11.043. PMID 15734365. S2CID 24988672.
- ^ Cheng Y, Yang CY, Lin CP, Lee PL, Decety J (May 2008). «Ауырсыну сезімі басқаларында соматосенсорлы тербелісті басады: магнитоэнцефалографиялық зерттеу». NeuroImage. 40 (4): 1833–40. дои:10.1016/j.neuroimage.2008.01.064. PMID 18353686. S2CID 1827514.
- ^ Моррисон, Мен .; Ллойд, Д .; Di Pellegrino, G.; Робертс, Н. (2004). "Vicarious responses to pain in anterior cingulate cortex: Is empathy a multisensory issue?". Когнитивті, аффективті және мінез-құлық неврологиясы. 4 (2): 270–8. дои:10.3758/CABN.4.2.270. PMID 15460933.
- ^ Wicker; т.б. (2003). "Neuron" (PDF). Архивтелген түпнұсқа (PDF) 2007-06-30.
- ^ Әнші; т.б. (2004). «Ғылым». Sciencemag.org. Журналға сілтеме жасау қажет
| журнал =(Көмектесіңдер) - ^ а б Jabbi, Mbemba; Swart, Marte; Keysers, Christian (2007). "Empathy for positive and negative emotions in the gustatory cortex". NeuroImage. 34 (4): 1744–53. дои:10.1016/j.neuroimage.2006.10.032. PMID 17175173. S2CID 13988152.
- ^ Lamm C, Batson CD, Decety J (January 2007). «Адам эмпатиясының жүйке субстраты: перспективалық және когнитивті бағалаудың әсерлері». Когнитивті неврология журналы. 19 (1): 42–58. дои:10.1162 / jocn.2007.19.1.42. PMID 17214562. S2CID 2828843.
- ^ Freedberg, David; Gallese, Vittorio (2007). "Motion, emotion and empathy in esthetic experience". Когнитивті ғылымдардың тенденциялары. 11 (5): 197–203. дои:10.1016 / j.tics.2007.02.003. PMID 17347026. S2CID 1996468.
- ^ Gazzola, Aziz-Zadeh and Keysers (2006). "Current Biology" (PDF). Архивтелген түпнұсқа (PDF) 2007-06-30.
- ^ Molenberghs; т.б. (2012 жылғы 2 қаңтар). "Activation patterns during action observation are modulated by context in mirror system areas" (PDF). NeuroImage. 59 (1): 608–15. дои:10.1016/j.neuroimage.2011.07.080. PMID 21840404. S2CID 13951700.
- ^ Ходжат, Мохаммадреза; т.б. "American Journal of Medical Quality | 28.1 (2013)" (PDF). 6-7 бет.
- ^ Hernandez-Lallement J, Attah AT, Soyman E, Pinhal CM, Gazzola V, Keysers C (2020). "Harm to Others Acts as a Negative Reinforcer in Rats". Curr Biol. 30 (6): 949–961.e7. дои:10.1016/j.cub.2020.01.017. PMID 32142701. S2CID 212424287.CS1 maint: бірнеше есімдер: авторлар тізімі (сілтеме)
- ^ Оберман, Л .; Рамачандран, В.С. (2009). «Айнадағы нейрондық жүйеге шағылысу: олардың эволюциялық функциялары моторды бейнелеуден тыс». Пинедада Дж.А. (ред.). Айна нейрондық жүйелер: Айна процестерінің әлеуметтік танымдағы рөлі. Humana Press. бет.39 –62. ISBN 978-1-934115-34-3.
- ^ Рамачандран, В.С. (1 қаңтар, 2009). «Өзін-өзі тану: соңғы шекара, Edge Foundation веб-очеркі». Алынған 26 шілде, 2011.
- ^ Skoyles, John R., Gesture, Language Origins, and Right Handedness, Psycholoqy: 11,#24, 2000
- ^ Petrides, Michael, Cadoret, Genevieve, Mackey, Scott (2005). Orofacial somatomotor responses in the macaque monkey homologue of Broca's area, Nature: 435,#1235
- ^ Porter Jr, R. J.; Lubker, J. F. (1980). "Rapid reproduction of vowel-vowel sequences: Evidence for a fast and direct acoustic-motoric linkage in speech". Сөйлеу және есту мәселелерін зерттеу журналы. 23 (3): 593–602. дои:10.1044/jshr.2303.593. PMID 7421161.
- ^ McCarthy, R.; Warrington, E. K. (1984). "A two-route model of speech production. Evidence from aphasia". Brain : A Journal of Neurology. 107 (2): 463–485. дои:10.1093/brain/107.2.463. PMID 6722512.
- ^ McCarthy, R. A.; Warrington, E. K. (2001). "Repeating Without Semantics: Surface Dysphasia?". Нейроказа. 7 (1): 77–87. дои:10.1093/neucas/7.1.77. PMID 11239078. S2CID 12988855.
- ^ Marslen-Wilson, W. (1973). «Тілдік құрылым және өте қысқа кешігулерде сөйлеу көлеңкесі». Табиғат. 244 (5417): 522–523. Бибкод:1973Natur.244..522M. дои:10.1038 / 244522a0. PMID 4621131. S2CID 4220775.
- ^ Fay, W. H.; Coleman, R. O. (1977). "A human sound transducer/reproducer: Temporal capabilities of a profoundly echolalic child". Ми және тіл. 4 (3): 396–402. дои:10.1016/0093-934X(77)90034-7. PMID 907878. S2CID 29492873.
- ^ Schippers, MB; Roebroeck, A; Renken, R; Нанетти, Л; Keysers, C (2010). "Mapping the Information flow from one brain to another during gestural communication" (PDF). Proc Natl Acad Sci U S A. 107 (20): 9388–93. Бибкод:2010PNAS..107.9388S. дои:10.1073/pnas.1001791107. PMC 2889063. PMID 20439736. Архивтелген түпнұсқа (PDF) 2012-02-27.
- ^ Моро, Андреа (2008). Вавилон шекаралары. Мүмкін емес тілдердің миы мен жұмбақтары. MIT түймесін басыңыз. б. 257. ISBN 978-0-262-13498-9.
- ^ Longo, M. R; Kosobud, A.; Bertenthal, B. I. (April 2008). "Automatic imitation of biomechanically possible and impossible actions: effects of priming movements versus goals" (PDF). J Exp Psychol Hum Percept орындау. 34 (2): 489–501. дои:10.1037/0096-1523.34.2.489. PMID 18377184.
- ^ Kosonogov, Vladimir (2011). "Listening to action-related sentences impairs postural control". Электромиография және кинезиология журналы. 21 (5): 742–745. дои:10.1016/j.jelekin.2011.05.007. ISSN 1050-6411. PMID 21705230.
- ^ Chartrand, T. L.; Bargh, J. A. (June 1999). "The chameleon effect: the perception-behavior link and social interaction". J Pers Soc Psychol. 76 (6): 893–910. дои:10.1037/0022-3514.76.6.893. PMID 10402679. S2CID 11818459.
- ^ Lakin, J. L.; Chartrand, T. L. (July 2003). "Using nonconscious behavioral mimicry to create affiliation and rapport". Психологиялық ғылым. 14 (4): 334–9. дои:10.1111/1467-9280.14481. PMID 12807406. S2CID 8458849.
- ^ van Baaren, R. B.; Maddux, W. W.; Chartrand, T. L.; De Bouter, C.; van Knippenberg, A. (May 2003). "It takes two to mimic: behavioral consequences of self-construals". Тұлға және әлеуметтік психология журналы. 84 (5): 1093–102. дои:10.1037/0022-3514.84.5.1093. PMID 12757151. S2CID 729948.
- ^ Heyes, Cecilia (2011). "Automatic imitation". Психологиялық бюллетень. 137 (3): 463–83. дои:10.1037/a0022288. PMID 21280938. S2CID 6975248.
- ^ Paukner A, Suomi SJ, Visalberghi E, Ferrari PF (August 2009). "Capuchin monkeys display affiliation toward humans who imitate them". Ғылым. 325 (5942): 880–3. Бибкод:2009Sci...325..880P. дои:10.1126/science.1176269. PMC 2764469. PMID 19679816.
- ^ а б Oberman LM, Hubbard EM, McCleery JP, Altschuler EL, Ramachandran VS, Pineda JA., EEG evidence for mirror neuron dysfunction in autism spectral disorders Мұрағатталды 2007-01-03 Wayback Machine, Brain Res Cogn Brain Res.; 24(2):190-8, 2005-06
- ^ Dapretto, M; Davies, MS; Pfeifer, JH; Scott, AA; Sigman, M; Bookheimer, SY; Iacoboni, M (2006). «Өзгелердегі эмоцияны түсіну: аутизм спектрі бұзылған балалардағы айна нейрондық дисфункциясы». Табиғат неврологиясы. 9 (1): 28–30. дои:10.1038 / nn1611. PMC 3713227. PMID 16327784.
- ^ Dapretto M, Davies MS, Pfeifer JH, et al. (Қаңтар 2006). «Өзгелердегі эмоцияны түсіну: аутизм спектрі бұзылған балалардағы айна нейрондық дисфункциясы». Нат. Нейросчи. 9 (1): 28–30. дои:10.1038 / nn1611. PMC 3713227. PMID 16327784.
- ^ Perkins T, Stokes M, McGillivray J, Bittar R (October 2010). "Mirror neuron dysfunction in autism spectrum disorders". Клиникалық неврология журналы. 17 (10): 1239–43. дои:10.1016/j.jocn.2010.01.026. PMID 20598548. S2CID 15141982.
- ^ Oberman LM, Ramachandran VS, Pineda JA (April 2008). "Modulation of mu suppression in children with autism spectrum disorders in response to familiar or unfamiliar stimuli: the mirror neuron hypothesis". Нейропсихология. 46 (5): 1558–65. дои:10.1016/j.neuropsychologia.2008.01.010. PMID 18304590. S2CID 14280719.
- ^ Churcland, P.S. (2011). «6». Braintrust. Принстон университетінің баспасы. б. 156.
- ^ Аутизмдегі әрекеттерді түсінудің екі жүйесі: айна және ментализация, Барон-Коэн, Тагер-Флусберг және Ломбардо, Басқа ойларды түсіну тарауы. [2]
- ^ Hadjikhani, N; т.б. (2006). "Anatomical Differences in the Mirror Neuron System and Social Cognition Network in Autism". Ми қыртысы. 16 (9): 1276–82. дои:10.1093/cercor/bhj069. PMID 16306324.
- ^ https://www.newscientist.com/article/dn18837-mirror-neurons-seen-behaving-normally-in-autism.html
- ^ Hamilton, A. F. (August 2009). "Goals, intentions and mental states: challenges for theories of autism" (PDF). Балалар психологиясы және психиатриясы журналы. 50 (8): 881–892. CiteSeerX 10.1.1.621.6275. дои:10.1111 / j.1469-7610.2009.02098.x. PMID 19508497.
- ^ Murphy, P; Brady, N; Фицджералд, М; Troje, NF (2009). "No evidence for impaired perception of biological motion in adults with autistic spectrum disorders". Нейропсихология. 47 (14): 3225–35. дои:10.1016/j.neuropsychologia.2009.07.026. PMID 19666038. S2CID 12495492.
- ^ Saygin, A.; Кук Дж .; Blakemore, S. (2010-10-18). Baker, Chris I (ed.). "Unaffected perceptual thresholds for biological and non-biological form-from-motion perception in autism spectrum conditions". PLOS One. 5 (10): e13491. Бибкод:2010PLoSO...513491S. дои:10.1371/journal.pone.0013491. PMC 2956672. PMID 20976151.
- ^ Cook, Jennifer; Сайгин, Айсе Пинар; Swain, Rachel; Blakemore, Sarah-Jayne (2009). "Reduced sensitivity to minimum-jerk biological motion in autism spectrum conditions". Нейропсихология. 47 (14): 3275–3278. дои:10.1016/j.neuropsychologia.2009.07.010. PMC 2779370. PMID 19632248.
- ^ Fan, Y. T.; Дети, Дж .; Yang, C. Y.; Liu, J. L.; Cheng, Y. (September 2010). "Unbroken mirror neurons in autism spectrum disorders". Балалар психологиясы және психиатриясы журналы. 51 (9): 981–988. дои:10.1111/j.1469-7610.2010.02269.x. PMID 20524939.
- ^ https://www.newscientist.com/article/dn18837-mirror-neurons-seen-behaving-normally-in-autism/
- ^ Gordon R (1986). "Folk psychology as simulation". Ақыл мен тіл. 1 (2): 158–171. дои:10.1111/j.1468-0017.1986.tb00324.x.
- ^ Goldman A. (1989). "Interpretation psychologized". Ақыл мен тіл. 4 (3): 161–185. дои:10.1111/j.1468-0017.1989.tb00249.x.
- ^ Meltzoff AN, Decety J (March 2003). "What imitation tells us about social cognition: a rapprochement between developmental psychology and cognitive neuroscience". Лондон Корольдік қоғамының философиялық операциялары. B сериясы, биологиялық ғылымдар. 358 (1431): 491–500. дои:10.1098 / rstb.2002.1261. PMC 1351349. PMID 12689375.
- ^ Sommerville JA, Decety J (April 2006). «Әлеуметтік өзара әрекеттестік тінін тоқу: моторлық таным аясындағы дамудың психологиясы мен когнитивті неврологиясын тұжырымдау». Психономдық бюллетень және шолу. 13 (2): 179–200. дои:10.3758 / BF03193831. PMID 16892982. S2CID 14689479.
- ^ Keysers C, Gazzola V (2007). "Integrating simulation and theory of mind: from self to social cognition" (PDF). Когнитивті ғылымдардың тенденциялары. 11 (5): 194–196. дои:10.1016 / j.tics.2007.02.002. PMID 17344090. S2CID 18930071. Архивтелген түпнұсқа (PDF) 2009-03-25.
- ^ Haroush K, Williams ZM (March 2015). "Neuronal prediction of opponent's behavior during cooperative social interchange in primates". Ұяшық. 160 (6): 1233–45. дои:10.1016/j.cell.2015.01.045. PMC 4364450. PMID 25728667.
- ^ Cheng YW, Tzeng OJ, Decety J, Imada T, Hsieh JC (July 2006). "Gender differences in the human mirror system: a magnetoencephalography study". NeuroReport. 17 (11): 1115–9. дои:10.1097 / 01.wnr.0000223393.59328.21. PMID 16837838. S2CID 18811017.
- ^ Cheng Y, Decety J, Lin CP, Hsieh JC, Hung D, Tzeng OJ (June 2007). "Sex differences in spinal excitability during observation of bipedal locomotion". NeuroReport. 18 (9): 887–90. дои:10.1097 / WNR.0b013e3280ebb486. PMID 17515795. S2CID 16295878.
- ^ Yang CY, Decety J, Lee S, Chen C, Cheng Y (January 2009). "Gender differences in the mu rhythm during empathy for pain: an electroencephalographic study". Миды зерттеу. 1251: 176–84. дои:10.1016 / j.brainres.2008.11.062. PMID 19083993. S2CID 40145972.
- ^ Cheng Y, Lee PL, Yang CY, Lin CP, Hung D, Decety J (May 2008). «Адамның айна-нейрондық жүйесінің му-ритміндегі гендерлік айырмашылықтар». PLOS ONE. 3 (5): e2113. Бибкод:2008PLoSO ... 3.2113C. дои:10.1371 / journal.pone.0002113. PMC 2361218. PMID 18461176.
- ^ Schulte-Rüther, M; Markowitsch, HJ; Шах, Ндж; Fink, GR; Piefke, M (Aug 1, 2008). "Gender differences in brain networks supporting empathy". NeuroImage. 42 (1): 393–403. дои:10.1016/j.neuroimage.2008.04.180. PMID 18514546. S2CID 10461927.
- ^ а б c Джалал, Баланд; Ramachandran, Vilayanur S. (2017). "Sleep Paralysis, "The Ghostly Bedroom Intruder" and Out-of-Body Experiences: The Role of Mirror Neurons". Адам неврологиясының шекаралары. 11: 92. дои:10.3389/fnhum.2017.00092. ISSN 1662-5161. PMC 5329044. PMID 28293186.
- ^ McCormick LM, Brumm MC, Beadle JN, Paradiso S, Yamada T, Andreasen N (March 2012). "Mirror neuron function, psychosis, and empathy in schizophrenia". Психиатрияны зерттеу. 201 (3): 233–9. дои:10.1016/j.pscychresns.2012.01.004. PMC 3545445. PMID 22510432.
Әрі қарай оқу
- Hickok, Greg (2014). The Myth of Mirror Neurons. ISBN 978-0-393-08961-5.
- Keysers, C. (2011). The Empathic Brain. ISBN 978-1-4637-6906-2.
- Iacoboni M, Mazziotta JC (2007). «Айна нейрондық жүйесі: негізгі нәтижелер және клиникалық қолдану». Энн Нейрол. 62 (3): 213–8. дои:10.1002 / ана.21198. PMID 17721988. S2CID 3225339.
- Keysers, C., & Gazzola, V. (2006), Towards a unifying neural theory of social cognition, Progress in Brain Research.[3]
- Morsella, E., Bargh, J.A., & Gollwitzer, P.M. (Eds.) (2009). Oxford Handbook of Human Action. Нью Йорк: Оксфорд университетінің баспасы.
- Preston, S. D.; де Ваал, Ф.Б.М. (2002). «Эмпатия: оның түпкілікті және жақын негіздері». Мінез-құлық және ми туралы ғылымдар. 25 (1): 1–72. дои:10.1017 / s0140525x02000018. PMID 12625087.
- Rizzolatti G, Fabbri-Destro M, Cattaneo L (2009). "Mirror neurons and their clinical relevance". Nat Clin Pract Neurol. 5 (1): 24–34. дои:10.1038/ncpneuro0990. PMID 19129788. S2CID 2979216.
- Rizzolatti, G., Sinigaglia, C. (2008). Mirrors in the Brain. How We Share our Actions and Emotions. Oxford (UK), Оксфорд университетінің баспасы.
Сыртқы сілтемелер
- NOVA scienceNOW: Mirror Neurons (including a 14-minute broadcast segment)
- You remind me of me in the New York Times.
- 7 Minute Video on TED.com :: Neuroscientist Vilayanur Ramachandran outlines the functions of mirror neurons
- Mirror Neuron Forum, Perspectives on Psychological Science, September, 2011
- Talking Brains, Greg Hickok and David Poeppel, News And Views On the Neural Organization of Language
- Thomas, Ben: What's So Special about Mirror Neurons? Scientific American Guest Blog, 2012 (an overview of prominent research approaches based on interviews with Iacoboni, Hickok, Heyes and Gallese)
- Mirror neurons: still an open question? Paolo B. Pascolo. Progress in Neuroscience 2013; 1 (1-4): 25-82