Solanaceae - Solanaceae - Wikipedia
| Solanaceae | |
|---|---|
 | |
| Гүлдену Brugmansia suaveolens бастап АҚШ ботаникалық бағы | |
| Ғылыми классификация | |
| Корольдігі: | Планта |
| Клайд: | Трахеофиттер |
| Клайд: | Ангиоспермдер |
| Клайд: | Eudicots |
| Клайд: | Астеридтер |
| Тапсырыс: | Соланалес |
| Отбасы: | Solanaceae Джусс. |
| Subfamilies[1] | |

The Solanaceae, немесе түнгі көлеңкелер, а отбасы туралы гүлді өсімдіктер бір жылдық және көпжылдық шөптерден жүзім сабақтарына дейін, лиана, эпифиттер, бұталар мен ағаштар жатады және олардың қатарына жатады ауыл шаруашылығы дақылдар, дәрілік өсімдіктер, дәмдеуіштер, арамшөптер және сәндік өсімдіктер. Отбасының көптеген мүшелерінде күшті заттар бар алкалоидтар, ал кейбіреулері өте жоғары улы, бірақ көп - соның ішінде қызанақ, картоп, баялды, қоңырау және Чили бұрышы Ретінде қолданылады тамақ. Отбасы бұйрыққа жатады Соланалес, ішінде астерид топ және сынып Магнолиопсида (қосжарнақтылар ).[2] Solanaceae 98-ге жуық және 2700 түрден тұрады,[3] үлкен әртүрлілікпен тіршілік ету ортасы, морфология және экология.
Solanaceae атауы тектен шыққан Соланум, «түнгі өсімдік». Этимологиясы Латын сөз түсініксіз. Бұл атау белгілі бір сортаң гүлдердің күн мен оның сәулелеріне ұқсастығынан туындауы мүмкін. Кем дегенде бір түрі Соланум «күнбағыс» деп аталады. Сонымен қатар, атау латынша етістіктен туындауы мүмкін solare, «тыныштандыру» деген мағынаны білдіреді, болжамды түрде тыныштандыруға қатысты фармакологиялық кейбір қасиеттері психоактивті отбасының түрлері.
Отбасының бүкіл әлем бойынша таралуы бар, олардан басқа барлық континенттерде бар Антарктида. Түрлердегі ең үлкен әртүрлілік Оңтүстік Америка және Орталық Америка. 2017 жылы ғалымдар а-ны ашқаны және талдағаны туралы хабарлады қазба томатилло табылған Патагония Аргентинаның 52 миллион жылға созылған аймағы Б.П. Табылу Solanaceae өсімдіктер тобының алғашқы көрінісін артқа тастады.[4] Томатилло басқа түнгі мезгілдерге қарағанда кеш дамығандықтан, бұл Solanaceae алғашқы пайда болған болуы мүмкін дегенді білдіруі мүмкін. Мезозой эрасы.[5]
Solanaceae бірқатар жиналған немесе өсірілетін түрлерді қамтиды. Отбасының экономикалық жағынан маңызды тұқымы[дәйексөз қажет ] болып табылады Соланум, құрамында ботташық (S. tuberosum, шын мәнінде, отбасының тағы бір жалпы атауы - «картоп отбасы»), қызанақ (S. lycopersicum), және баялды немесе баклажан (S. melongena). Тағы бір маңызды түр, Capsicum, екеуін де шығарады Чили бұрышы және болгар бұрышы.
Тұқым Physalis деп аталатын жер шабақтарын шығарады, сонымен қатар томатилло (Physalis philadelphica), Мүйізді қарлыған және Қытай шамы. Тұқым Лиций Босткорндар мен қасқыр Лиций барбарумы. Никотиана басқа түрлерімен қатар, темекі. Соланасеяның кейбір басқа маңызды мүшелеріне бірқатар сәндік өсімдіктер жатады Петуния, Browallia, және Личианттар, және психоактивті алкалоидтардың қайнар көздері, Датура, Мандрагора (mandrake), және Atropa belladonna (өлім түні). Кейбір түрлері дәрілік мақсатта қолданылуымен, психотроптық әсерімен немесе улы болуымен кеңінен танымал.
Экономикалық маңызды тұқымдастардың көп бөлігі субфамилияда бар Соланоидтар, темекіні қоспағанда (Nicotiana tabacum, Nicotianoideae) және петуния (Петуния × гибрида, Petunioideae).
Темекі және петуния сияқты көптеген соланацеялар қолданылады модельді организмдер кезінде негізгі биологиялық сұрақтарды зерттеу кезінде ұялы, молекулалық, және генетикалық деңгейлер.
Этимология және айтылу
«Solanaceae» атауы (АҚШ: /ˌсoʊлəˈneɪсмен,-сменˌaɪ,-сменˌeɪ,-сменˌмен/) келеді халықаралық ғылыми лексика бастап Жаңа латын, бастап Соланум, типтегі түр, + -жасушалар,[6] стандартталған жұрнақ қазіргі таксономиядағы өсімдік тегі үшін. Тұқым атауы Классикалық латын сөз солон түнгі түндерге қатысты (әсіресе Solanum nigrum ), «мүмкін сол, 'күн', + -анум, бейтарап -анус."[6]
Сипаттама
Бұл бөлім үшін қосымша дәйексөздер қажет тексеру. (Сәуір 2015) (Бұл шаблон хабарламасын қалай және қашан жою керектігін біліп алыңыз) |

Соланацеядағы өсімдіктер шөптер түрінде болуы мүмкін, бұталар, ағаштар, жүзім және лиана, кейде эпифиттер. Олар болуы мүмкін жылдық, екіжылдықтар, немесе көпжылдық өсімдіктер, тік немесе декументті. Кейбіреулерінде жерасты бар түйнектер. Оларда жоқ кешіктірушілер, не латекс және түсті емес сабс. Олар жапырақтардың базальды немесе түпкілікті тобына ие болуы мүмкін немесе осы түрлердің ешқайсысында болмайды. The жапырақтары жалпыға қарсы немесе ауыспалы болып келеді (яғни өсімдіктің түбінде кезектеседі және қарсыға қарама-қарсы) гүлшоғыры ). Жапырақтары шөпті, былғары немесе айналуы мүмкін тікенектер. Жапырақтары жалпы петиолат немесе жерасты, сирек отырықшы. Олар жиі жағымсыз, бірақ кейбіреулері хош иісті немесе ұрықсыз. Жапырақты ламина қарапайым да, күрделі де болуы мүмкін, ал соңғысы пиннатифидті немесе тернаталы болуы мүмкін. Жапырақтары торлы венацияға ие және базальды емес меристема. Ламиналар әдетте дорсивентральды және секреторлы қуыстарға ие емес. The стоматалар әдетте жапырақтың екі жағының бірімен шектеледі; олар екі жақта да сирек кездеседі.
The гүлдер негізінен гермафродиттер, бірақ кейбіреулері бар біртұтас, andromonoecious, немесе екі қабатты түрлері (мысалы, кейбіреулері) Соланум немесе Симонантус). Тозаңдану энтомофильді. Гүлдер жалғыз немесе терминалды, цимоза немесе қолтық асты гүлшоғырына топтастырылуы мүмкін. Гүлдері орташа, хош иісті (Никотиана), фетид (Антоцерцис) немесе жағымсыз. Гүлдер әдетте актиноморфты, сәл зигоморфты, немесе айқын зигоморфты (мысалы, билабиалды королла гүлдерінде Шизантус түрлер). Симметриядағы бұзушылықтар байланысты болуы мүмкін андроциум, дейін периант, немесе екеуі де бір уақытта. Түрлердің басым көпшілігінде гүлдерде тостағаншасы бар сараланған периант болады королла (сәйкесінше бес сеппальмен және бес жапырақшамен) бес кастрөлден тұратын андроциум стаменс және екі кілемдер қалыптастыру гинеций бастықпен аналық без[7] (сондықтан олар пентамерлер және тетрациклді деп аталады). The стаменс болып табылады эпипетальды және әдетте төрт немесе бес, көбінесе төрт немесе сегізге көбейеді. Әдетте оларда гипогинді диск бар. Тостағаншаның гамосепальды түрі бар (сепальдар бір-біріне қосылып, түтік түзеді), (4) 5 (6) сегменттері тең, оның бес үлесі бар, қабықшалары түтікке қарағанда қысқа, ол тұрақты және жиі акцентрлі. Короллада әдетте бес жапырақ бар, олар түтікті біріктіреді. Гүл формалары әдетте айналмалы (дөңгелек тәрізді, бір жазықтықта жайылып, қысқа түтікшемен) немесе түтікшелі (ұзартылған цилиндрлік түтік), кампануляцияланған немесе шұңқыр тәрізді болады.
Андроцейдің ішінде (2) (4) 5 (6) бос ерлер бар, оппозицепалдар (яғни олар жапырақшалармен кезектеседі), олар көбінесе құнарлы немесе кейбір жағдайларда (мысалы, Salpiglossideae) оларда стаминодтар болады. Екінші жағдайда, әдетте, бір стаминод бар (Сальпиглоссис) немесе үш (Шизантус). Антериялар олардың жоғарғы жағына сақина түзеді немесе олар толығымен бос, дорсификсирленген немесе порицидтік дегисценциямен немесе ұсақ бойлық жарықтар арқылы базификстелген. Ерлер жіп пішінді немесе жалпақ болуы мүмкін. Таяқшаларды кораллин түтігінің ішіне кіргізуге немесе енгізуге болады. Өсімдіктер бір мезгілде микроспорогенез көрсетеді, микроспоралар - тетрада, тетраэдр немесе изобилат. Тозаң түйіршіктері дегисценция сәтінде екіжасушалы, әдетте ашық және бұрыштық болады.
Гиноциум қосарпелярлы (сирек үш немесе бес локалды) жоғары аналық без және екі локулалар, ол екіншіден жалғанға бөлінуі мүмкін септа, Nicandreae және Datureae үшін жағдай. Гинеоций гүлдің медианалық жазықтығына қатысты қиғаш жағдайда орналасқан. Оларда бар стиль және бір стигма; соңғысы қарапайым немесе билобатты. Әрбір локуляцияда акиллярлы плацентациясы бар анатропты немесе гемианатропты болып табылатын бір-ден 50-ге дейін овуля бар. Дамыту эмбрион қап сияқты болуы мүмкін Көпбұрыш немесе Аллиум түрлері. Эмбрион қаптары ядролық полюстер бұрын еріген ұрықтандыру. Үш антипод әдетте жағдайдағыдай уақытша немесе тұрақты болады Atropa. The жеміс болуы мүмкін жидек қызанақ немесе қасқыр сияқты жағдайдағыдай а дегисцент капсула сияқты Датуранемесе а дрюпа. Жемісі бар осьтік плацентация. Капсулалар қалыпты жағдайда септицидті немесе сирек локулицидті немесе вальватты болады. The тұқымдар әдетте эндоспермиялық, майлы (сирек крахмал) және айқын түктері жоқ. Көптеген соланацеялардың тұқымдары дөңгелек және тегіс, диаметрі шамамен 2-4 мм (0,079-0,157 дюйм). Эмбрион түзу немесе қисық болуы мүмкін және екі котиледоны бар. Solanaceae түрлерінің көпшілігінде 2n = 24 болады хромосомалар,[8] бірақ бұл сан 12-ге үлкен еселік болуы мүмкін полиплоидия. Жабайы картоп, оның 200-ге жуығы, көбінесе диплоидты (2 × 12 = 24 хромосома), бірақ триплоидты (3 × 12 = 36 хромосома), тетраплоид (4 × 12 = 48 хромосома), пентаплоид (5 × 12 = 60) және тіпті гексаплоидты (6 × 12 = 72 хромосома) түрлер немесе популяциялар бар. Мәдени түрлер Solanum tuberosum 4 × 12 = 48 хромосома бар. Кейбіреулер Capsicum түрлерінде 2 × 12 = 24 хромосома, ал басқаларында 26 хромосома бар.
Сипаттамалардың әртүрлілігі
Алдыңғы сипаттамаға қарамастан, Solanaceae репродуктивті сипаттамаларында да үлкен морфологиялық өзгергіштікті көрсетеді. Бұл әртүрліліктің мысалдары:[9][10]
- Гиноциумды құрайтын карпельдер саны
Жалпы, Solanaceae-де екі карпельден түзілген гиноеций (гүлдің аналық бөлігі) болады. Алайда, Меланантус монокарпелярлы гиноциумы бар, ішінде үш-төрт кілем бар Capsicum, үштен беске дейін Никандра, кейбір түрлері Джабороза және Трианаеа және төрт кілем Iochroma umbellatum.
- Аналық бездегі локулалар саны
Аналық бездегі локулалар саны, әдетте, тұтастардың санымен бірдей. Алайда, кейбір түрлері жалған септа (әр локуланы бөлетін ішкі қабырғалар) болуына байланысты сандар бірдей болмайтын түрлер пайда болады, мысалы Датура және Lycieae-нің кейбір өкілдері (тұқымдастар) Грабовския және Вассобия).
- Жұмыртқа жасушаларының типі және олардың саны
Жұмыртқалар негізінен төңкерілген, күрт артқа бүктелген (анатропты), бірақ кейбір тұқымдастарда сабақтарына (кампилотропты) тік бұрышпен айналатын жұмыртқалар болады. Фродус, Грабовския немесе Вассобия) немесе ішінара төңкерілген (сияқты гемитропты) Цеструм, Capsicum, Шизантус және Лиций). Әрбір локулаға шәует саны бірнешеден өзгереді (әр локулада екі жұп Грабовския, әрбір локалда бір жұп Лиций) және кейде локалда тек бір ғана жұмыртқа болады, мысалы Меланантус.
- Жемістердің түрі
Solanaceae-нің басым көпшілігінің жемістері - жидектер немесе капсулалар (соның ішінде пирксидия), ал сирек друптер. Жидектер Cestroideae, Solanoideae субфамилияларында кең таралған (қоспағанда) Датура, Ориктус, Грабовския және Hyoscyameae тайпасы) және Juanulloideae тайпасы (қоспағанда Маркеа ). Капсулалар Cestroideae субфамилияларына тән (қоспағанда Цеструм ) және Schizanthoideae, Salpiglossoideae және Anthocercidoideae тайпалары және туысы Датура. Hyoscyameae тайпасында пирксидия бар. Drupes Lycieae тайпасына және иохроминаларға тән.[11]
Алкалоидтар
Алкалоидтар ретінде өсімдіктер шығаратын азотты органикалық заттар болып табылады екінші метаболит және жануарларға өте төмен физиологиялық әсер ететін, тіпті төмен дозада.[дәйексөз қажет ] Solanaceae алкалоидтардың әртүрлі диапазонына ие екендігі белгілі. Адамдар үшін бұл алкалоидтар қалаулы, улы немесе екеуі де болуы мүмкін. The тропандар Соланасеяда кездесетін алкалоидтардың ішіндегі ең танымал болып табылады. Құрамында осы заттар бар өсімдіктер ғасырлар бойы улану ретінде қолданылып келеді. Алайда, улар деп танылғанына қарамастан, бұл заттардың көпшілігі баға жетпес фармацевтикалық қасиеттерге ие. Көптеген түрлер құрамында аз-кем белсенді немесе улы болуы мүмкін әртүрлі алкалоидтар бар скополамин, атропин, гиосциамин, және никотин. Олар хенбан сияқты өсімдіктерде кездеседі (Hyoscyamus albus), белладонна (Atropa belladonna ), джимсон арамшөптері (Драмалық страмониум ), мандрэйк (Mandragora autumnalis ), темекі және басқалары. Алкалоидтардың кейбір негізгі түрлері:
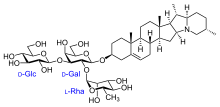
- Соланин: A улы гликоалкалоид ащы дәмімен оның С формуласы бар45H73ЖОҚ15. Оны соланидин алкалоиды а түзеді көмірсу бүйір тізбек. Ол картоп пен қызанақ сияқты жапырақтарда, жемістерде және әр түрлі соланастардың түйнектерінде кездеседі. Оның өндірісі шөп қоректенетін жануарларға қарсы адаптивті қорғаныс стратегиясы деп саналады. Интоксикация соланиннен асқазан-ішек жолдарының бұзылуымен сипатталады (диарея, құсу, іштің ауыруы) және жүйке аурулары (галлюцинация және бас ауруы ). The өлімге әкелетін орташа доза дене салмағының 2 мен 5 мг / кг аралығында. Симптомдар ішке қабылдағаннан кейін 8-12 сағаттан кейін көрінеді. Картоптағы бұл гликоалкалоидтардың мөлшері, мысалы, оларды өсіру кезіндегі қоршаған орта жағдайына, сақтау ұзақтығына және сортына байланысты айтарлықтай өзгереді. Гликоалкалоидтың орташа концентрациясы - 0,075 мг / г картоп.[12] Соланин кейде жидектерді жеген адамдардағы уланулар үшін жауапты болды Solanum nigrum немесе Solanum dulcamara немесе жасыл картоп.[13][14]

- Tropanes: «Тропан» термині а түр олар табылған, Atropa ( белладонна тұқымдас). Atropa атымен аталады Грек тағдыры, Атропос, кім өмірдің жібін кесіп тастады. Бұл номенклатура оның уыттылығы мен өлімдігін көрсетеді. Олар бициклді органикалық азот қосылыстары (IUPAC номенклатура: 8-метил-8-азабицикло [3.2.1] октан), химиялық формуласы С8H15N. Бұл алкалоидтарға, басқалармен қатар, атропин, кокаин, скополамин, және гиосциамин. Олар әртүрлі түрлерде кездеседі, мысалы, мандраге (Mandragora officinarum және M. autumnalis ), қара хенбэн немесе сасық түнгі бояу (Hyoscyamus niger ), белладонна (Atropa belladonna ), джимсон арамшөпі немесе шайтанның тұзағы (Драмалық страмониум ) және Бругмансия , сондай-ақ Solanaceae отбасындағы көптеген басқа адамдар.[15] Фармакологиялық тұрғыдан, олар ең танымал болып табылады антихолинергия болмыста, яғни олар тежейді неврологиялық арқылы берілетін сигналдар эндогендік нейротрансмиттер, ацетилхолин. Көбінесе олар көптеген түрлерін тоқтата алады аллергиялық реакциялар. Артық дозалану белгілері болуы мүмкін құрғақ ауз, кеңейтілген оқушылар, атаксия, зәрді ұстау, галлюцинация, құрысулар, кома және өлім. Әдетте қолданылатын атропин офтальмологиялық агент, қарашықтарды кеңейтеді және осылайша көздің ішкі жағын тексеруді жеңілдетеді. Шындығында, жидектерден алынған шырын A. belladonna кезінде итальяндық сыпайылар қолданған Ренессанс оқушыларының кеңеюін туғызу арқылы олардың көздерінің мөлшерін көбейту. Тропанның улылығына қарамастан, олар өте аз мөлшерде енгізілгенде пайдалы дәрілік заттар болып табылады. Олар өзгерте алады холинергиялық улану, бұл органофосфаттың шамадан тыс әсерінен болуы мүмкін инсектицидтер және химиялық соғыс сияқты агенттер зарин және VX. Скополамин (табылған Hyoscyamus muticus және Scopolia carniolica ) ретінде қолданылады құсуға қарсы қарсы теңіз ауруы немесе зардап шегетін адамдарға арналған жүрек айну алу нәтижесінде химиотерапия.[16][17] Скополамин мен гиосциамин - парасимпатикалық жүйке жүйесіне әсер етуіне байланысты фармакологияда және медицинада ең көп қолданылатын тропан алкалоидтары. Атропиннің а стимулятор әсер етеді орталық жүйке жүйесі және жүрек, ал скополаминнің а седативті әсер. Бұл алкалоидтарды басқа кез келген класс қосылыстары алмастыра алмайды, сондықтан олар әлі де сұранысқа ие. Бұл алкалоидтардың, қатысатын ферменттердің және оларды өндіретін гендердің метаболизмін зерттеудің белсенді өрісін дамытудың себептерінің бірі. Мысалы, 6-β-гидроксилаза гиосциамині, тропанның биосинтетикалық жолының соңында скополамин өндірісіне әкелетін гиосциаминнің гидроксилденуін катализдейді. Бұл фермент оқшауланған және сәйкес ген үш түрден клондалған: H. нигер, A. belladonna және B. candida.[18][19][20]

- Никотин: Никотин (IUPAC номенклатура (S) -3- (1-метилпирролидин-2-ыл) пиридин) а пирролидин -де көп мөлшерде өндірілген алкалоид темекі өсімдік (Nicotiana tabacum). Баклажан, қызанақ, картоп, бұрыш сияқты жеуге жарамды соланецтердің құрамында никотин де бар, бірақ концентрациялары темекіден 100000-1000000 есе аз.[21][22] Өсімдіктегі никотиннің қызметі қорғаныс қызметін атқарады шөп қоректілер, өйткені бұл өте тиімді нейротоксин, атап айтқанда қарсы жәндіктер. Шындығында, никотин көптеген жылдар бойы ан ретінде қолданылып келеді инсектицид дегенмен, оны қолдану қазіргі кезде оның құрылымынан алынған синтетикалық молекулалармен ауыстырылуда. Төмен концентрацияда никотин сүтқоректілерде стимулятор рөлін атқарады, бұл темекі шегушілерге тәуелділікті тудырады. Тропан тәрізді, ол холинергиялық нейрондарға әсер етеді, бірақ керісінше әсер етеді (бұл ан агонист қарама-қарсы антагонист ). Оның жоғары ерекшелігі бар никотиндік ацетилхолинді рецепторлар басқа ACh ақуыздарына қарағанда.
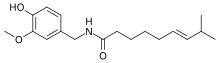
- Капсаицин: Капсаицин (IUPAC номенклатура 8-метил-N-ваниллил-транс-6-нонамид) никотин мен тропандардан құрылымдық жағынан ерекшеленеді. Ол тұқымдас түрлерінде кездеседі Capsicum қамтиды чилис және хабанерос және оны анықтайтын белсенді ингредиент Scoville рейтингі осы дәмдеуіштердің Қосылыс адамдар үшін айтарлықтай улы емес. Алайда, бұл сүтқоректілердің көпшілігінде ерекше ауыру рецепторларын ынталандырады, әсіресе жылуды қабылдаумен байланысты ауыз қуысының шырышты қабаты және басқа да эпителий ұлпалары. Капсаицин осы шырышты қабаттармен байланысқа түскенде, ол өрттен туындаған күйіктен өзгеше жану сезімін тудырады. Капсаицин құстарға емес, тек сүтқоректілерге әсер етеді. Бұрыш тұқымы құстардың ас қорыту жолдарынан аман қала алады; олардың тұқымдары өніп-жетілуге жетілгеннен кейін олардың жемістері ашық түсті болады, сол арқылы тұқымдарды тарататын құстардың назарын аударады. Капсаицин сығындысы жасау үшін қолданылады бұрыш спрейі, агрессивті және бейбіт сүтқоректілерге қарсы пайдалы тежеу.
Тарату

Соланасеяның мүшелері барлық жерде кездеседі континенттер Антарктидадан басқа түрлердің ең көп түрлілігі кездеседі Орталық Америка және Оңтүстік Америка. Әртүрлілік орталықтары да кездеседі Австралия және Африка. Solanaceae көптеген басқа түрлерді алады экожүйелер, бастап шөлдер дейін тропикалық ормандар және көбінесе бұзылған аймақтарды колониялайтын екінші реттік өсімдіктерде кездеседі. Жалпы бұл тұқымдастағы өсімдіктер тропикалық және қоңыржай таралған.
Зауыт иесі
Картоп түйнегі көбелегі (Фторимаея оперкулелла ) - олигофагты жәндік, ол Solanaceae тұқымдасының өсімдіктерімен, әсіресе картоп өсімдігімен қоректенуді жөн көреді (Solanum tuberosum). Әйел P. operculella жапырақтарды жұмыртқалау үшін пайдаланыңыз, ал жұмыртқадан шыққан личинкалар жапырақтың мезофиллін жейді. Жапырақтарды тамақтандырғаннан кейін, личинкалар сүңгіп, өсімдіктің түйнектері мен тамырларымен қоректенеді.[23]
Таксономия
Соланасейлердің, соның ішінде субфамилияларды, тайпалар мен тұқымдарды қосқандағы келесі таксономиялық синопсис ең соңғы молекулалық филогенетика отбасын зерттеу:[2][3][24][25]
Cestroideae (Browallioideae)




Бұл субфамилия перициклді талшықтардың болуымен сипатталады, төрт немесе бес стаменді андроциум, жиі диаминамозды. Хромосоманың негізгі сандары өте өзгермелі, х = 7-ден х = 13-ке дейін. Субфамилия сегіз тұқымдас (үш тайпаға бөлінген) және бүкіл Америкада таралған 195-ке жуық түрден тұрады. Тұқым Цеструм ең маңызды болып табылады, өйткені оның құрамында субфамилиядағы 195 түрдің 175 түрі бар. The Cestreae тайпа ерекше, өйткені оның құрамына ұзын хромосомалары бар таксондар кіреді (ұзындығы 7,21-ден 11,511 мкм-ге дейін), қалған отбасы әдетте қысқа хромосомаларға ие (мысалы, никотианоидайда 1,5 пен 3,52 мкм аралығында).
- Browallieae Хунц.
- Browallia Л., таралған алты түрі бар тұқым Неотропикалық аймақ дейін Аризона ішінде АҚШ
- Стрептозолен Miers, Анды мекендейтін монотипті туыс
- Cestreae тайпасы Дон, ағаш тұқымды өсімдіктердің үш тұқымы, бұталар
- Salpiglossideae тайпасы (Бесінші. ) Хунц.
- Рейезия Гей, төрт түрі, үшеуі Чилидің солтүстігінде, ал біреуі солтүстік Чилиде және солтүстік Аргентинада.
- Сальпиглоссис Руис және Пав., Оңтүстік Америкадан шыққан екі түр
Гетцеоидтер


Бұл кіші отбасы жемісті және ұрық тәрізді ұрықтардың, қисық эмбриондармен және ірі етті котиледондардың болуымен сипатталады. Негізгі хромосома саны х = 13. Оған төрт тұқымдастар мен бес түр кіреді Үлкен Антиль аралдары. Кейбір авторлар өздерінің молекулалық деректері монотиптік текті көрсетеді деп болжайды Цоала Боссер & D'Arcy эндемикалық болып табылатын осы семьяға қосу керек Мадагаскар, және Metternichia оңтүстік-шығысында Бразилия. Goetzeaceae Airy Shaw осы подфамилияның синонимі ретінде қарастырылады.[26]
- Coeloneurum Радлк., монотиптік эндемик Испаниола
- Эспадаеа Rchb., монотипті, бастап Куба
- Гетцеа Видлер, Антиль аралдарынан екі түрді қамтиды
- Хенуния Грисеб., монотипті, шыққан Куба
Никотианоидалар

- Антоцерцидтер Г.Дон: Австралияда эндемик болған бұл тайпа жеті тұқымда 31 түрден тұрады. Рудың молекулярлық филогенетикалық зерттеулері оның қарындасы екенін көрсетеді Никотиана, және тұқымдастар Антоцерцис, Антотрош, Граммосолен, және Симонантус болып табылады монофилетикалық. Кейбір сипаттамалар тайпаның ішінен шыққан деп есептейді, мысалы, жартылай шеңберлі оперкуламен, брактеолат гүлдерімен және жемістер ретінде жидектермен бір көзді шыңдар.[27]
- Антоцерцис Лабиль., 10 түрі, Австралия
- Антотрох Endl., төрт түрі, Австралия
- Кренидиум Хэги, монотипті түр, Австралия
- Цифертера Miers, 9 түрі, Австралия
- Duboisia Р.Бр., төрт түрі, Австралия
- Граммосолен Хэги, екі түрі, Австралия
- Симонантус Хэги, екі түрі, Австралия
- Nicotianeae тайпасы Дум.
- Никотиана Л., 52 американдық, 23 австралиялық және бір африкалық түрімен таралған тұқым
Petunioideae



Молекулалық филогенетика Petunioideae-дің апалы-сіңілі екенін көрсетеді қаптау х = 12 хромосомасы бар субфамилиялардың (Соланоидтар және Никотианоидалар ). Олардың құрамында калистегиндер, тропандарға ұқсас алкалоидтар бар. Андроциум төрт ұзыннан тұрады (сирек бес), әдетте екі түрлі ұзындыққа ие. Бұл субфамилияның негізгі хромосома саны x = 7, 8, 9 немесе 11 болуы мүмкін. Ол Орталық және Оңтүстік Америкада таралған 13 тұқымдас пен 160-қа жуық түрден тұрады. Молекулалық деректер тұқым Патагонияда шыққан деп болжайды. Бентамиелла, Комбера, және Пантацанта Goetzeoideae тұқымдасына жататын тайпа (Benthamielleae) қатарына жатқызуға болатын клад жасаңыз.
- Бентамиелла Speg., Патагониядан шыққан 12 түр
- Бушетия Дунал, үш неотропикалық түр
- Брунфелсия Л., неотропиктен 45 түрге жуық
- Калибрахоа Церв. бұрынғы La Llave & Лекс., неотропикадан 32 түрден тұрады. Морфологиялық мәліметтер бұл түрдің құрамына кіру керектігін ұсынады Петуния. Алайда, молекулалық және цитогенетикалық деректер екеуін де бөлек ұстау керектігін көрсетеді. Шынында, Калибрахоа х = 9 негізгі хромосома саны бар, ал ол Петуния х = 7.[28][29]
- Комбера Сэндв., Патагониядан шыққан екі түр
- Фабиана Руис және Пав., Анды мекендейтін 15 түр
- Хунзикерия D'Arcy, АҚШ-тың оңтүстік-батысынан үш түр және Мексика
- Лептоглоссис Бесінші., батыс Оңтүстік Америкадан жеті түр
- Ниерембергия Руис және Пав., Оңтүстік Америкадан 21 түр
- Пантацанта Speg., Патагониядан шыққан моноспецификалық түр
- Петуния (Джусс. ) Виджман, Оңтүстік Америкадан 18 түр
- Пломания Хунц. & Subils, Мексика мен Гватемаладан шыққан монотипті тұқым
Шизантоида

Schizanthoideae-ге бір жылдық және екі жылдық тропан алкалоидтары бар, перициклді талшықтары жоқ, шаштары мен тозаң дәндері бар өсімдіктер. Гүлдері зигоморфты. Андроциумда екі стамен және үш стаминод бар, тозаңсыздандыру жарылғыш болып табылады. Жеміс түрі бойынша шизантоида тұқымдасын сақтайды плезиоморфты Solanaceae тұқымдасының жеміс формасы, капсулалар, an анемохоралық, дисперсияның абиотикалық түрі. Бұл Schizanthoidae-де ерте дивергенцияның генетикалық шектеулеріне байланысты (төменде қараңыз), сондай-ақ шизантус эволюциясы және ашық мекенде болу.[30]Эмбрион қисық. Негізгі хромосома саны х = 10. Шизантус қатты зигоморфты гүлдері мен негізгі хромосомалар санына байланысты соланасейлер арасында біршама атиптік тұқым. Морфологиялық және молекулалық мәліметтерден көрінеді Шизантус басқа соланасеяларға қарындас тұқымдас және басқалардан ерте, ал кейінірек бөлініп кеткен Бор немесе басында Кайнозой, 50 миллион жыл бұрын.[24][25] Ішіндегі гүл түрлерінің алуан түрлілігі Шизантус бұл түрлердің Жерорта теңізінде, биік альпілік және шөл экожүйелерінде болған, содан кейін Чилиде және Аргентинаның көршілес аймақтарында болған әр түрлі тозаңдатқыштардың түрлеріне бейімделуінің өнімі болды.[31]
- Шизантус Руис және Пав., Чилиден шыққан 12 түр.
Швенкиоидтер
Перициклді талшықтары бар біржылдық өсімдіктер, олардың гүлдері зигоморфты, андроецийде төрт динамикалық штамм немесе үш стаминод бар; эмбрион түзу және қысқа. Негізгі хромосома саны х = 12. Оған Оңтүстік Америкада таралған төрт тұқым мен 30-ға жуық түр кіреді.
- Гетерантия Нис & Наурыз., бір түрі Бразилиядан
- Меланантус Walp., Бразилия, Куба және Гватемаладан алынған бес түр
- Протошвенкия Soler , Боливия мен Бразилиядан шыққан монотипті тұқым, кейбір молекулярлық филогенетикалық зерттеулер бұл тұқымның субфамилияда белгісіз таксономиялық жағдайға ие екендігін болжады
- Швенкия Л., Американың неотропикалық аймақтарына таралған 22 түрі
Соланоидтар













- Capsiceae Дюморт
- Capsicum Л. қабылданған 40 неотропикалық түрді қамтиды[32]
- Личианттар (Дунал) Хасслер, бүкіл Америкада және Азияда таралған 200 түр
- Табиғат Г.Дон, екі тұқым морфологиялық және молекулалық деңгейде керемет ажыратылады, Бругмансия ағаш түрлерін қамтиды, ал Датура құрамында шөптер немесе бұталар бар, соңғы тұқымды үш бөлімге бөлуге болады: Страмониум, Дутра және Ceratocaulis.[33] Монотиптік түр Тромпетия жақында Боливия бұтағын бұрын орналастыру үшін орналастырылды Iochroma cardenasianum - қазір Datureae-ге тиесілі және бұрын ойлағандай Physaleae емес.[34]
- Бругмансия Жеке тұлға, Анд тауларынан алты түр
- Датура Л., 12 неотропикалық түр
- Тромпетия Дж.Дупин, Андтан шыққан жалғыз түрлер Боливия
- Гиосиямдар Endl.
- Анисод Сілтеме, төрт түрі Қытай, Үндістан және Гималай
- Архихоскиям А.М.Лу, Түркия мен Иранның жалғыз түрлері.
- Atropa Л., төрт еуроазиялық түр[32]
- Атропант Пасчер, Қытайдан шыққан монотипті тұқым
- Hyoscyamus Л., Қабылданған 10 түр[32] Жерорта теңізінен Қытайға дейін таратылды
- Фисхлея Г.Дон, 6 қабылданған еуроазиялық түрлер[32]
- Пржевальск Максим., бір түрі Қытайдан
- Скополия Жак., екі еуропалық және екі түрден бөлінген Шығыс Азия.
- Джаборозия Miers
- Джабороза Джусс., Оңтүстік Американың 23 түрін қамтитын тұқым.
- Соландрея Miers
- Juanulloinae субтрибі неотропикалық таралуы бар 10 тұқымдас ағаштар мен эпифитті бұталардан тұрады.[35] Осы тұқымдардың кейбіреулері (Диссохрома, Merinthopodium және Трианаеа) түрлеріне айқын тәуелділікті көрсетеді жарқанаттар тұқымдарды тозаңдандыру үшін де, дисперсиялау үшін де.[36]
- Диссохрома Miers, Бразилияның оңтүстігінен екі түр
- Эктозома Miers
- Хоккесиофитон Хунц.
- Хуануллоа Руис және Пав., Оңтүстік және Орталық Америкадан 11 түр
- Маркеа Бай., Оңтүстік және Орталық Америкадан 9 түр
- Merinthopodium Дж. Донн. Sm. Оңтүстік Америкадан шыққан үш түр
- Раховориана Д 'Арси
- Шултесиантус Хунц., сегіз неотропикалық түр
- Трианаеа Планш. & Линден, Оңтүстік Американың алты түрі
- Subtribe Solandrinae, монотипті субтрианның, Хуануллинадан айырмашылығы, оның эмбриондарында қазіргі жұмыс істейтін котилондар мен жартылай төменгі аналық бездер болады.[35]
- Соландра Sw., Американың неотропикалық аймақтарынан 10 түр
- Juanulloinae субтрибі неотропикалық таралуы бар 10 тұқымдас ағаштар мен эпифитті бұталардан тұрады.[35] Осы тұқымдардың кейбіреулері (Диссохрома, Merinthopodium және Трианаеа) түрлеріне айқын тәуелділікті көрсетеді жарқанаттар тұқымдарды тозаңдандыру үшін де, дисперсиялау үшін де.[36]
- Lycieae Хунц. құрғақ немесе жартылай құрғақ климатта өсетін ағаш тұқымды өсімдіктердің үш тұқымдасы бар. Космополит Лиций тайпадағы ең көне және морфологиялық өзгергіштікке ие.[37] Молекулалық филогенетикалық зерттеулер екеуін де ұсынады Грабовския және Фродус ішіне қосу керек Лиций,[38] және осы тұқым, бірге Нолана және Склерофилакс, қазіргі уақытта таксономиялық санаты жоқ кладты (Lyciina) құрайды.[26] Құстар таратқан қызыл етді жидектер жемістердің негізгі түрі болып табылады Лиций. Бұл тектегі жемістердің әр түрлі түрлері жаңа айтылған жидек түрінен тұқымдары азайтылған друпаға айналды.[39]
- Грабовския Schltdl., Оңтүстік Америкадан үш түр
- Лиций Л., 83 космополит түрі
- Фродус Miers, Чилидің солтүстігінде эндемик болатын екі түрі
- Мандрагорея (Веттст.) Хунц. & Барбоза молекулалық филогенетикалық зерттеулерге сәйкес тайпаның белгілі жүйелік позициясы жоқ.[26]
- Мандрагора Л., Еуразиядан шыққан екі түр
- Никандрия Wettst. Оңтүстік Американың екі тегі бар тайпа. Молекулярлық филогенетикалық зерттеулер тұқымдастардың бір-бірімен байланысы жоқтығын және олардың басқа тұқымдастармен байланысы жоқ екенін көрсетеді, сондықтан олардың таксономиялық жағдайы белгісіз.[26]
- Exodeconus Раф., Оңтүстік Американың алты түрі
- Никандра Аданс, бір түрі неотропикалық аймақтарға таралған
- Ноланеа Rchb. көбінесе шырынды жапырақтары бар шөптер мен ұсақ бұталар, олар ақтан түрлі-түсті көгілдір реңктерге дейін өте әдемі гүлдері бар, олардың жемістері шизокарпалы, әр түрлі жаңғақтарды тудырады.
- Нолана Л., 89 түрі Оңтүстік Американың батысында таралған
- Физалия Miers, Capsiceae қарындасы болып табылатын үлкен тайпа.
- Iochrominae жазылымы (Miers) Hunz., Physaleae тайпасының ішіндегі қаптама. құрамында алты түрге бөлінген, негізінен Анд тауларында таралған 37 түр бар. Бұл қосылыстың мүшелері ағаш тәрізді бұталар немесе тартымды түтікшелермен немесе бұрылған гүлдермен ұсақ ағаштармен сипатталады. Олар сондай-ақ отбасында бар барлық түрлерден тұратын үлкен гүлді әртүрлілікке ие. Олардың гүлдері қызыл, қызғылт сары, сары, жасыл, көк, күлгін немесе ақ түсті болуы мүмкін. Королла айналуы үшін түтікшелі болуы мүмкін, әр түрлі түрлер арасындағы түтік ұзындығы сегіз есеге дейін өзгереді.[40]
- Физалина (Miers) Hunz. , монофилді субтрит, 10 тұқымдасты қамтиды және оған аралармен тозаңданған сары, ақ немесе күлгін жалғыз аксиларлы гүлдері бар шөптер немесе ағаш бұталар жатады. Тозаңдану пайда болғаннан кейін, королла түсіп, тостағанша дамып келе жатқан сиқырды толығымен жапқанша кеңейеді (тостаған акцесцент деп аталады). Көптеген түрлерде тостаған жетілген кезде сары немесе сарғыш түске боялады. Жидектерде жасыл немесе сары-сарғыш тұқымдар көп, көбінесе қызыл немесе күлгін түске боялған.[41]
- Брахист Miers, Мексикадан және Орталық Америкадан үш түр
- Хамаесарача (А.Грей) Бесінші. & Ілмек., Мексикадан және Орталық Америкадан 10 түрі бар.
- Дарциантус, тек Боливия мен Перуден шыққан 1 түрге жататын тұқым.
- Лейкофизалис Ридберг, Америка Құрама Штаттары мен Мексиканың оңтүстік батысынан 3 түрді қамтиды.
- Маргарантус Шлехт., Мексикадан 1 түрімен.
- Oryctes Уотсон, АҚШ-тың оңтүстік батысынан монотипті тұқым.
- Physalis Л., Американың тропикалық аймақтары арқылы таралған 85 түрімен және 1 түрімен таралатын ең үлкен тұқымдас Қытай.
- Хинкула Раф. тек АҚШ-тың оңтүстік батысы мен Мексикадан 1 түрімен.
- Целталия, тұқымдас Physalis, 2 түрімен бүкіл Мексикаға таралған және Гватемала.
- Ветерингия L 'Heritier, неотропикалық аймақтардан 15 түрі бар тұқым.
- Subpribe Salpichroinae, бұл Physaleae тұқымы, оған 1 тұқымда таралған 16 американдық түр кіреді:
- Нектуция Кунт., Мексикаға тән монотипті тұқым.
- Сальпичроа Miers, Андардан және Оңтүстік Американың басқа аймақтарынан 15 түрі бар тұқым.
- Subanet Withaninae - бұл таралуы кең таралатын Physaleae, оның 9 тұқымы:
- Архифизалис Куанг, Қытай мен Жапониядан келген 3 түрімен.
- Афина Сендн., оған Бразилиядан 7 түр кіреді.
- Аврелиана Сендн., Оңтүстік Америкадан 5 түрімен.
- Куатрезия Хунц., 11 неотропикалық түрімен. Молекулалық зерттеулер бұл түрдің бірге екендігін көрсетеді Депреа және Ларнакс белгісіз таксономиялық жағдайға ие.[26]
- Депреа Раф., 6 неотропикалық түрімен.
- Ларнакс Miers, көптеген таксономистер оны синоним деп санайды Депреа, Анды мекендейтін 22 түрден тұрады.
- Меллисия Ілмек. f., монотиптік түр Әулие Елена жалпы атауы бар Әулие Хелена ағашы (жуырда түрге енген Withania)
- Nothocestrum А.Грей
- Physaliastrum Макино, 10 азиат түрімен (жуырда түрге енген тұқым Withania).
- Tubocapsicum (Wettst.) Макино, тек Қытайда кездесетін бір түрімен.
- Withania Паук., оның 10 түрі бар Канар аралдары, Африка және Непал.
- Solaneae тайпасы. Тұқым Цифомандра Сендн., Дископодиум Хохст., Нормания Лоу, Тригуера Кав. және Lycopersicum Диірмен аударылды Соланум. Субтитр екі тұқымнан тұрады:[26]
- Джалтомата Schltdl.құрамында 50 неотропикалық түр бар.
- Соланум Л., бүкіл әлем бойынша 1328 түр таралған, ең үлкен тұқымдас ангиоспермалардың бірі.
Incertae sedis

Төмендегі тұқымдар әлі күнге дейін соланецтер ішіндегі танылған субфамилияларға орналастырылмаған (incertae sedis ).
- Duckeodendron Кульманнб, монотипті түр Амазонка тропикалық орманы.
- Парабушетия Байлон, аз танымал, монотипті түр Бразилия.
- Пауия Деб. & Дутта, монотиптік түр Ассам және Аруначал-Прадеш Н.Е.Үндістан
Түрлердің таралуы және таралуы

Solanaceae 98 тұқымдас және 2700 түрден тұрады. Түрлердің бұл өте бай болуына қарамастан, олар тұқымдастар арасында біркелкі бөлінбейді. Төмендегі кестеде көрсетілгендей, ең маңызды сегіз тұқымдастың түрлерінің 60% -дан астамы бар. Соланум - отбасын типтейтін тұқым - соланасеяның жалпы түрлерінің 50% құрайды.
| Ұрпақ | Түрлердің шамамен саны |
|---|---|
| Соланум | 1,330 |
| Личианттар | 200 |
| Цеструм | 150 |
| Нолана | 89 |
| Physalis | 85 |
| Лиций | 85 |
| Никотиана | 76 |
| Брунфелсия | 45 |
| Отбасындағы түрлердің болжамды саны | 2,700 |
Экономикалық маңызы



Solanaceae тұқымдасында картоп сияқты маңызды тамақ түрлері бар (Solanum tuberosum ), қызанақ (Solanum lycopersicum ), бұрыш (Capsicum annuum ) және баклажан немесе жұмыртқа өсімдігі (Solanum melongena ). Nicotiana tabacum, Бастапқыда Оңтүстік Америкадан шыққан, қазір бүкіл әлемде темекі өндіру үшін өсіріледі, көптеген сортаңдар әлемнің әртүрлі бөліктеріндегі маңызды арамшөптер болып табылады. Олардың маңыздылығы қоздырғыштарды немесе мәдени өсімдіктердің ауруларын орналастыра алатындығында, сондықтан олардың болуы өнімнің жоғалуын немесе жиналған өнімнің сапасын жоғарылатады. Бұған мысал келтіруге болады Acnistus arborescens және Browalia americana сол жүргізуші трипс байланысты мәдени өсімдіктерге зиян келтіреді,[42] және белгілі бір түрлері Датура кейінірек мәдени сортаңға берілетін вирустың әр түріне иелік етеді.[43] Сияқты арамшөптердің кейбір түрлері, Solanum mauritianum Оңтүстік Африкада жәндіктерді пайдалану арқылы биологиялық бақылау жасау мақсатында зерттеулер жүргізіліп жатқан осындай күрделі экологиялық және экономикалық проблемалар бар.[44]
Solanaceae-ге жататын өсімдіктердің алуан түрлері және олардың сорттары сәндік ағаштар, бұталар, бір жылдық және шөпті көпжылдықтар түрінде өсіріледі.[45] Мысалдарға мыналар жатады Brugmansia x candida («Періштенің трубасы») труба тәрізді үлкен маятник гүлдері үшін өсірілген немесе Brunfelsia latifolia, оның гүлдері өте хош иісті және 3 күн ішінде түсін күлгіннен аққа өзгертеді. Өздерінің тартымды гүлдері үшін өсірілетін бұталардың басқа түрлері Lycianthes rantonnetii (Көк картоп бұтасы немесе Парагвай түнгі көлеңкесі) күлгін көк гүлдермен және Nicotiana glauca («Ағаш темекісі») Сәндік өсімдік ретінде өсетін басқа сортаң тұқымдар мен тұқымдар петуния (Петуния × гибрида), Лиций, Соланум, Цеструм, Калибрахоа × буданы және Соландра. Арасында гибрид те бар Петуния және Калибрахоа (бұл жаңаны құрайды нотогенді емес деп аталады × Петчоа Г.Бокер және Дж. Шоу) сәндік заттар ретінде сатылады.[46][47] Көптеген басқа түрлер, атап айтқанда алкалоидтар шығаратындар фармакологияда және медицинада қолданылады (Никотиана, Hyoscyamus, және Датура ).
Solanaceae және геном
Осы отбасына жататын көптеген түрлер, олардың ішінде темекі мен қызанақ бар модельді организмдер биологиялық мәселелерді зерттеу үшін қолданылатын. Соланасея аспектілерінің бірі геномика is an international project that is trying to understand how the same collection of genes and proteins can give rise to a group of organisms that are so morphologically and ecologically different. The first objective of this project was to sequence the геном of the tomato. In order to achieve this each of the 12 хромосомалар of the tomato’s haploid genome was assigned to different sequencing centres in different countries. So chromosomes 1 and 10 were sequenced in the United States, 3 and 11 in China, 2 in Корея, 4 in Britain, 5 in India, 7 in France, 8 in Japan, 9 in Spain and 12 in Italy. The sequencing of the митохондриялық genome was carried out in Argentina and the хлоропласт genome was sequenced in the Еуропа Одағы.[48][49]
Сондай-ақ қараңыз
Әдебиеттер тізімі
- ^ "Solanaceae Juss., nom. минус ». Germplasm Resources ақпараттық желісі. Америка Құрама Штаттарының Ауыл шаруашылығы министрлігі. 2007-04-12. Алынған 2009-04-16.
- ^ а б Олмстед, Р.Г .; Sweere, J. A.; Spangler, R. E.; Bohs, L.; Palmer, J. D. (1999). "Phylogeny and provisional classification of the Solanaceae based on chloroplast DNA" (PDF). In Nee, M.; Symon, D. E.; Lester, R. N.; Jessop, J. P. (eds.). Solanaceae IV: advances in biology and utilization. The Royal Botanic Gardens. pp. 111–37.
- ^ а б Olmstead, R.G.; Bohs, L. (2007). "A Summary of molecular systematic research in Solanaceae: 1982-2006". Acta Horticulturae. 745 (745): 255–68. CiteSeerX 10.1.1.561.2269. дои:10.17660/ActaHortic.2007.745.11.
- ^ "Eocene lantern fruits from Гондванан Patagonia and the early origins of Solanaceae", Wilf et al, Ғылым, 06 Jan 2017, Vol. 355, Issue 6320, pp. 71-75, DOI: 10.1126/science.aag2737
- ^ "52 Million-Year-Old Tomatillo Fossils Rewrite Veggie History". NPR.org. Алынған 2017-01-20.
- ^ а б Merriam-Webster, Мерриам-Вебстердің сөздіксіз сөздігі, Merriam-Webster.
- ^ Yasin J. Nasir. "Solanaceae". Пәкістан флорасы.
- ^ Fujii, Kenjiro (1934). Cytologia. Botanical Institute. б. 281.
- ^ Hunziker, A.T. 1979: South American Solanaceae: a synoptic review. In: D'ARCY, W.G., 1979: The Biology and Taxonomy of the Solanaceae. Линн. Soc. Симптом. Сер. 7: p 48-85. Linnean Soc. & Academic Press; Лондон.
- ^ Balken, J.A. THE PLANT FAMILY SOLANACEAE:FRUITS IN SOLANACEAE «Мұрағатталған көшірме». Архивтелген түпнұсқа 2016-03-09. Алынған 2013-08-09.CS1 maint: тақырып ретінде мұрағатталған көшірме (сілтеме)
- ^ Армандо Т. Хунцикер: Соланацейлер тұқымдасы. A.R.G. Gantner Verlag KG, Ruggell, Лихтенштейн 2001. ISBN 3-904144-77-4
- ^ Zeiger, E. 1998. Solanine and Chaconine. Review of Toxicological Literature. Integrated Laboratory Systems, USA.«Мұрағатталған көшірме» (PDF). Архивтелген түпнұсқа (PDF) 2011-11-15. Алынған 2011-11-15.CS1 maint: тақырып ретінде мұрағатталған көшірме (сілтеме)
- ^ "Solanine poisoning". Br Med J. 2 (6203): 1458–9. 1979-12-08. дои:10.1136/bmj.2.6203.1458-a. PMC 1597169. PMID 526812.
- ^ Alexander RF, Forbes GB, Hawkins ES (1948-09-11). "A Fatal Case of Solanine Poisoning". Br Med J. 2 (4575): 518. дои:10.1136/bmj.2.4575.518. PMC 2091497. PMID 18881287.
- ^ Гриффин В.Ж., Лин Г.Д. (наурыз 2000). «Тропан алкалоидтарының химотаксономиясы және географиялық таралуы». Фитохимия. 53 (6): 623–37. дои:10.1016 / S0031-9422 (99) 00475-6. PMID 10746874.
- ^ Sneden, A. The tropane alkaloids. Medicinal Chemistry and Drug Design. Вирджиния достастығы университеті «Мұрағатталған көшірме» (PDF). Архивтелген түпнұсқа (PDF) 2007-09-27. Алынған 2007-09-27.CS1 maint: тақырып ретінде мұрағатталған көшірме (сілтеме)[сенімсіз ақпарат көзі ме? ]
- ^ Evans, W.C. 1979. Tropane alkaloids of the Solanaceae. En: HAWKES, LESTER and SHELDING (eds.). The biology and taxonomy of the Solanaceae. Линн. Soc. Симптом. Сер. 7:241-254. Linnean Soc. & Academic Press., London.
- ^ Matsuda, Jun; Okabe, Souichi; Хашимото, Такаси; Yamada, Yasuyuki (1991). "Molecular cloning of hyoscyamine 6β-hydroxylase, a 2-oxoglutarate-dependent dioxygenase, from cultured roots of Hyoscyamus niger". Биологиялық химия журналы. 266 (15): 9460–4. PMID 2033047.
- ^ Роча, Педро; Stenzel, Olaf; Парр, Адриан; Walton, Nicholas; Christou, Paul; Драгер, Биргит; Leech, Mark J (June 2002). "Functional expression of tropinone reductase I (trI) and hyoscyamine-6β-hydroxylase (сағ6сағ) бастап Hyoscyamus niger жылы Nicotiana tabacum". Өсімдік туралы ғылым. 162 (6): 905–13. дои:10.1016/S0168-9452(02)00033-X.
- ^ Cardillo, Alejandra B.; Giulietti, Ana M.; Marconi, Patricia L. (June 2006). "Analysis and sequencing of h6hmRNA, last enzyme in the tropane alkaloids pathway from anthers and hairy root cultures of Brugmansia candida (Solanaceae)". Биотехнологияның электронды журналы. 9 (3). дои:10.2225/vol9-issue3-fulltext-15.
- ^ Siegmund, Barbara; Leitner, Erich; Pfannhauser, Werner (1999-07-23). "Determination of the Nicotine Content of Various Edible Nightshades (Solanaceae) and Their Products and Estimation of the Associated Dietary Nicotine Intake". Дж. Агрик. Азық-түлік химиясы. 47 (8): 3113–3120. дои:10.1021/jf990089w. PMID 10552617. Алынған 2017-04-25.
- ^ Moldoveanu, Serban C.; Scott, Wayne A.; Lawson, Darlene M. (2016-04-01). "Nicotine Analysis in Several Non-Tobacco Plant Materials". Beiträge zur Tabakforschung International/Contributions to Tobacco Research. 27 (2): 54–59. дои:10.1515/cttr-2016-0008. ISSN 1612-9237. S2CID 155096089. Алынған 2017-05-05.
- ^ Varela, L. G.; Bernays, E. A. (1988-07-01). "Behavior of newly hatched potato tuber moth larvae,Phthorimaea operculella Zell. (Lepidoptera: Gelechiidae), in relation to their host plants". Жәндіктердің мінез-құлық журналы. 1 (3): 261–275. дои:10.1007/BF01054525. ISSN 0892-7553. S2CID 19062069.
- ^ а б Олмстед, Ричард Г. Palmer, Jeffrey D. (1992). "A Chloroplast DNA Phylogeny of the Solanaceae: Subfamilial Relationships and Character Evolution". Миссури ботаникалық бағының жылнамалары. 79 (2): 346–60. дои:10.2307/2399773. JSTOR 2399773.
- ^ а б Martins, Talline R.; Barkman, Todd J. (2005). "Reconstruction of Solanaceae Phylogeny Using the Nuclear Gene SAMT". Жүйелі ботаника. 30 (2): 435–47. дои:10.1600/0363644054223675. JSTOR 25064071. S2CID 85679774.
- ^ а б c г. e f Olmstead, R.G.; Bohs, L. (2007). "A Summary of molecular systematic research in Solanaceae: 1982-2006". Acta Horticulturae. 745 (745): 255–68. CiteSeerX 10.1.1.561.2269. дои:10.17660/ActaHortic.2007.745.11.
- ^ Garcia, Vicente F.; Olmstead, Richard G. (June 2003). "Phylogenetics of Tribe Anthocercideae (Solanaceae) Based on nhhF және трнL/F Sequence Data". Жүйелі ботаника. 28 (3): 609–15. дои:10.1043/02-52.1 (белсенді емес 2020-11-07). JSTOR 25063900.CS1 maint: DOI 2020 жылдың қарашасындағы жағдай бойынша белсенді емес (сілтеме)
- ^ Ando, Toshio; Kokubun, Hisashi; Watanabe, Hitoshi; Tanaka, Norio; Юкава, Томохиса; Hashimoto, Goro; Marchesi, Eduardo; Suárez, Enrique; Basualdo, Isabel L. (2005). "Phylogenetic Analysis of Petunia sensu Jussieu (Solanaceae) using Chloroplast DNA RFLP". Ботаника шежіресі. 96 (2): 289–97. дои:10.1093/aob/mci177. PMC 4246877. PMID 15944177.
- ^ Mishiba, Kei-Ichiro; Ando, Toshio; Mii, Masahiro; Watanabe, Hitoshi; Kokubun, Hisashi; Hashimoto, Goro; Marchesi, Eduardo (2000). "Nuclear DNA Content as an Index Character Discriminating Taxa in the Genus Petunia sensu Jussieu (Solanaceae)". Ботаника шежіресі. 85 (5): 665–73. дои:10.1006/anbo.2000.1122.
- ^ Knapp, S (2002). "Tobacco to tomatoes: a phylogenetic perspective on fruit diversity in the Solanaceae". Тәжірибелік ботаника журналы. 53 (377): 2001–2022. дои:10.1093/jxb/erf068. PMID 12324525.
- ^ Перес, Фернанда; Arroyo, Mary T. K.; Medel, Rodrigo; Hershkovitz, Mark A. (2006). "Ancestral reconstruction of flower morphology and pollination systems in Шизантус (Solanaceae)". Американдық ботаника журналы. 93 (7): 1029–38. дои:10.3732/ajb.93.7.1029. PMID 21642168.
- ^ а б c г. «Өсімдіктер тізімі, Atropa". Royal Botanic Garden, Kew.
- ^ Mace, E. S.; Gebhardt, C. G.; Lester, R. N. (1999). "AFLP analysis of genetic relationships in the tribe Datureae (Solanaceae)". Теориялық және қолданбалы генетика. 99 (3–4): 634–41. дои:10.1007/s001220051278. PMID 22665199. S2CID 30782826.
- ^ Дупин, Джулия; Smith, Stacey D (2017). "Phylogenetics of Datureae (Solanaceae), including description of the new genus Trompettia and re-circumscription of the tribe". Таксон. 67 (2): 359–375. дои:10.12705/672.6.
- ^ а б Кнапп, Сандра; Persson, Viveca; Blackmore, Stephen (1997). "A Phylogenetic Conspectus of the Tribe Juanulloeae (Solanaceae)". Миссури ботаникалық бағының жылнамалары. 84 (1): 67–89. дои:10.2307/2399954. JSTOR 2399954.
- ^ Sazima, M.; Buzato, S; Sazima, I (2003). "Dyssochroma viridiflorum (Solanaceae): A Reproductively Bat-dependent Epiphyte from the Atlantic Rainforest in Brazil". Ботаника шежіресі. 92 (5): 725–30. дои:10.1093/aob/mcg190. PMC 4244854. PMID 14500325.
- ^ Bernardello, Luis M. (1987). "Comparative Floral Morphology in Lycieae (Solanaceae)". Бриттония. 39 (1): 112–29. дои:10.2307/2806983. JSTOR 2806983. S2CID 84167107.
- ^ Levin, achel A.; Mille, Jill S. (2005). "Relationships within tribe Lycieae (Solanaceae): Paraphyly of Lycium and multiple origins of gender dimorphism". Американдық ботаника журналы. 92 (12): 2044–53. дои:10.3732/ajb.92.12.2044. JSTOR 4125537. PMID 21646122.
- ^ Bernardello, L.; Chiang-Cabrera, F. (1998). "A cladistic study on the American species of Лиций (Solanaceae) based on morphological variation". In Fortunato, Renée H; Bacigalupo, Nélida M (eds.). Proceedings of the VI Congreso Latinoamericano de Botánica, Mar del Plata, Argentina, 2-8 October, 1994. Миссури ботаникалық бағынан алынған жүйелі ботаникадағы монографиялар. Миссури ботаникалық бағының баспасы. бет.33–46. ISBN 978-0-915279-58-6.
- ^ Smith, Stacey DeWitt; Baum, David A. (2006). «Флоренетикалық әр түрлі Анды класының Iochrominae (Solanaceae) жабыны». Американдық ботаника журналы. 93 (8): 1140–53. дои:10.3732 / ajb.93.8.1140. JSTOR 4122802. PMID 21642180.
- ^ Whitson, Maggie; Manos, Paul S. (2005). "Untangling Physalis (Solanaceae) from the Physaloids: A Two-Gene Phylogeny of the Physalinae". Жүйелі ботаника. 30 (1): 216–30. дои:10.1600/0363644053661841. JSTOR 25064051. S2CID 86411770.
- ^ Masis, C. & Madrigal, R. 1994. Lista preliminar de malezas hospedantes de Thrips(Thysanoptera) que dañan al Хризантема морифолиумы en el valle central de Costa Rica. Agronomía Costarricense 18(1): 99-101. 1994 ж
- ^ Ormeño, J., Sepúlveda R., Rojas, R. Malezas del género Датура como factor epidemiológico del virus del mosaico de la alfalfa (amv), virus del mosaico del pepino (cmv) y virus y de la papa (pvy) en Solanáceas cultivadas. Agricultura técnica Vol. 66, Nº. 4, 2006, 333-341 Summary in Spanish
- ^ Pedrosa-Macedo, J., Olckers, T. & Vitorino, M. 2003. Phytophagous arthropods associated with Solanum mauritianum Scopoli (Solanaceae) in the first Plateau of Paraná, Brazil: a cooperative project on biological control of weeds between Brazil and South Africa. Неотроп. Энтомол. 32: 519-522. Article in English, with a summary in Portuguese
- ^ Arboles ornamentales cultivados en España. Solanáceas
- ^ Shaw, J. 2007. A new hybrid genus for Калибрахоа × Петуния (Solanaceae). HANBURYANA 2: 50–51 [1][тұрақты өлі сілтеме ]
- ^ The Value of Growing Petchoa SuperCal®. Ornamental News Oct 25 2012
- ^ International Tomato Sequencing Project Home
- ^ International Solanaceae Genomics Project (SOL), Systems Approach to Diversity and Adaptation.
- D'Arcy, William G. (1986). Solanaceae. Колумбия университетінің баспасы. ISBN 978-0-231-05780-6.
- Stevens, P. F. (2001–2007). "Solanaceae". Angiosperm филогенезінің веб-сайты, version 8, June 2007. Алынған 2007-11-04.
- Уотсон, Л .; Dallwitz, M. J. "Solanaceae". Гүлді өсімдіктердің тұқымдастары: сипаттамалар, иллюстрациялар, сәйкестендіру және ақпаратты іздеу. Version: 1 June 2007. Алынған 2007-11-04.
- Dimitri, M. 1987. Enciclopedia Argentina de Agricultura y Jardinería. Tomo I. Descripción de plantas cultivadas. Editorial ACME S.A.C.I., Buenos Aires.
- "Solanaceae Source". Алынған 2007-11-17.
- Hunziker, Armando T. 2001. The Genera of Solanaceae. A.R.G. Gantner Verlag K.G., Ruggell, Liechtenstein. ISBN 3-904144-77-4.
Әрі қарай оқу
- Хоукс, Дж. Г .; Lester, R. N.; Skelding, A. D. (1979). Соланацейлер биологиясы және таксономиясы. Academic Press, Лондон. ISBN 978-0-12-333150-2.
- Д'Арси, Уильям Г. (1986). Solanacea. Колумбия университетінің баспасы. ISBN 978-0-231-05780-6.
- Radford, Albert E. (1986). Fundamentals of Plant Systematics. Harper & Row, Publishers, Inc. ISBN 978-0-06-045305-3.
- Kubo, Ken-ichi; Paape, Timothy; Hatakeyama, Masaomi; Entani, Tetsuyuki; Takara, Akie; Kajihara, Kie; Tsukahara, Mai; Shimizu-Inatsugi, Rie; Shimizu, Kentaro K.; Takayama, Seiji (2015). "Gene duplication and genetic exchange drive the evolution of S-RNase-based self-incompatibility in Петуния" (PDF). Табиғат өсімдіктері. 1: 14005. дои:10.1038/nplants.2014.5. PMID 27246052. Түйіндеме – EurekAlert! (8 қаңтар, 2015).
Сыртқы сілтемелер
- Sol Genomics Network
- Solanaceae Network - pictures of plants
- Solanaceae көзі - A worldwide taxonomic monograph of all species in the genus Solanum.
- Solanaceae of Chile, by Chileflora[тұрақты өлі сілтеме ]
- Solanaceae жылы Л. Уотсон және М.Д. Даллвитц (1992 жылдан бастап). Гүлді өсімдіктердің тұқымдастары: сипаттама, иллюстрация, сәйкестендіру, ақпаратты іздеу. http://delta-intkey.com
- Solanaceae in USDA Plants Database.
- Family Solanaceae Израильдегі гүлдер
- SOL Genomics Network, Universidad de Cornell
- Imagines de various species of Solanaceae
- Solanaceae de Chile, by Chileflora
- Chilli: La especia del Nuevo Mundo (Article in Spanish by Germán Octavio López Riquelme regarding the biology, nutrition, culture and medical aspects of Chile.
- Solanaceae Resources on the Web
- Jäpelt RB, Jakobsen J (2013) Vitamin D in plants: a review of occurrence, analysis, and biosynthesis. Front Plant Sci 4, No. 136 -- Note the reference to higher cholesterol levels (and consequent Vitamin D3 levels) in family Solanaceae